

Tandem E2F binding sites in the promoter of the p107 cell cycle regulator control p107 expression and its cellular functions
- PMID: 20585628
- PMCID: PMC2891812
- DOI: 10.1371/journal.pgen.1001003
Tandem E2F binding sites in the promoter of the p107 cell cycle regulator control p107 expression and its cellular functions
Abstract
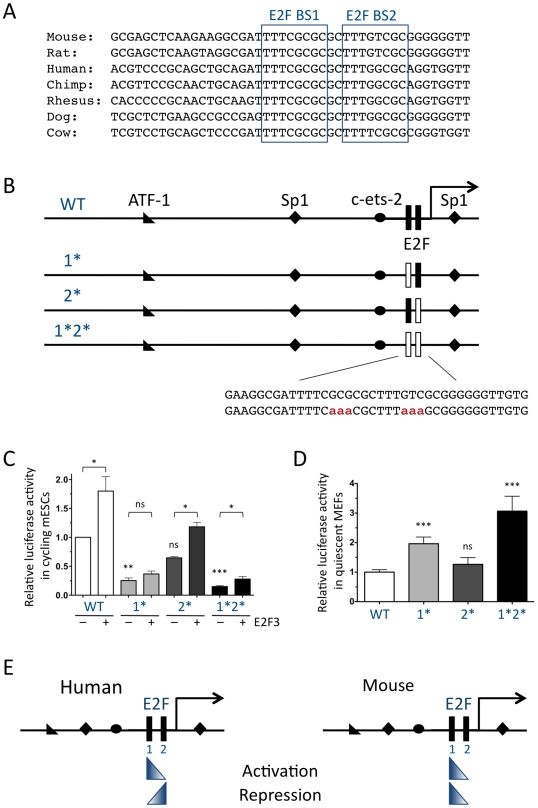
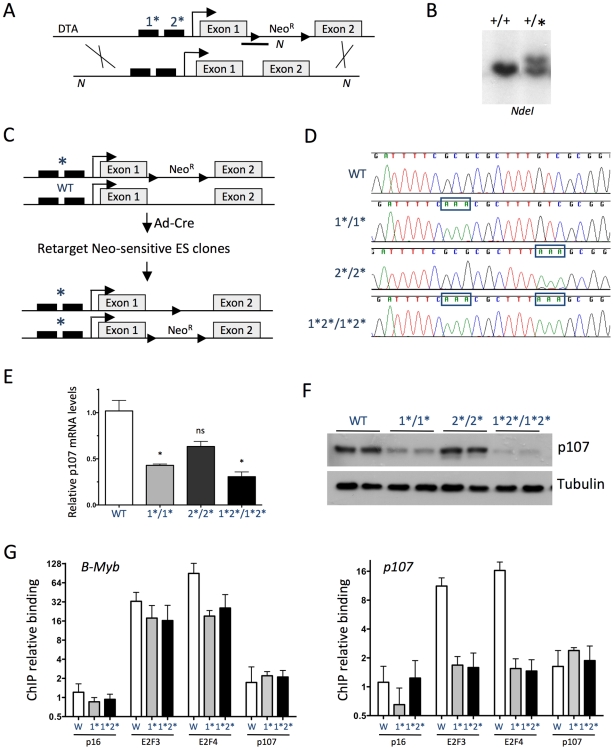
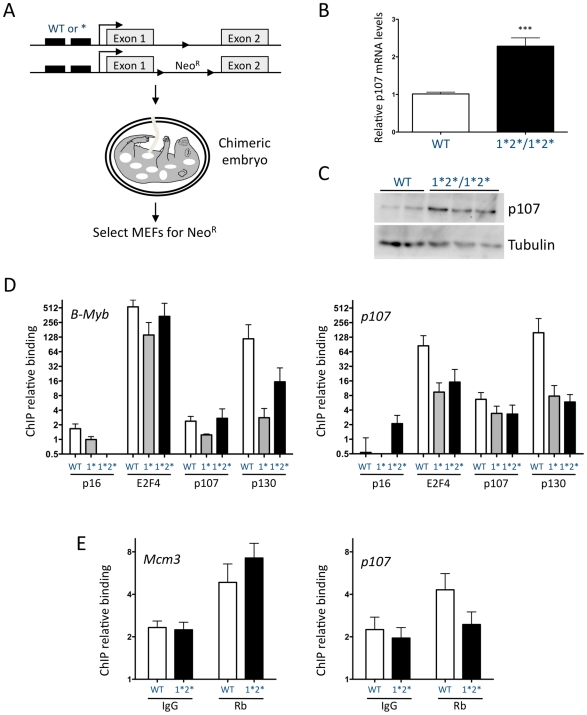
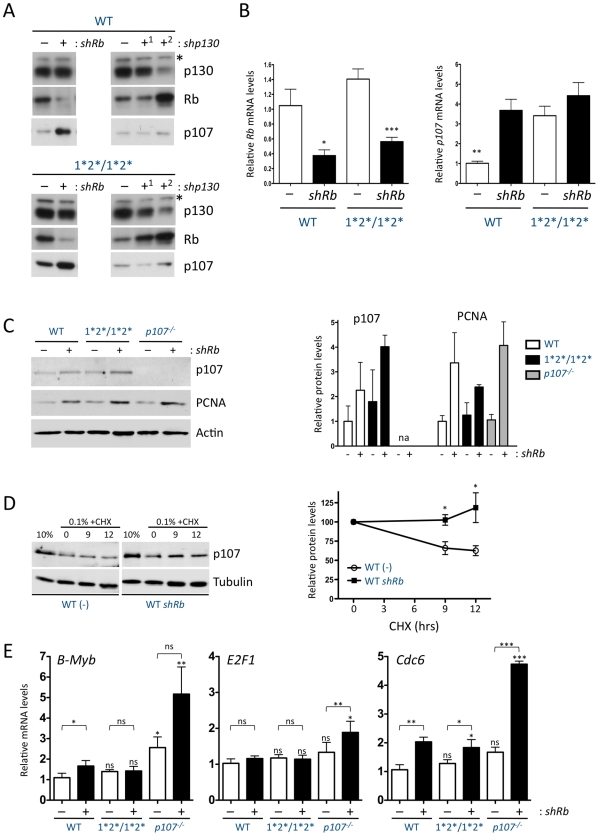
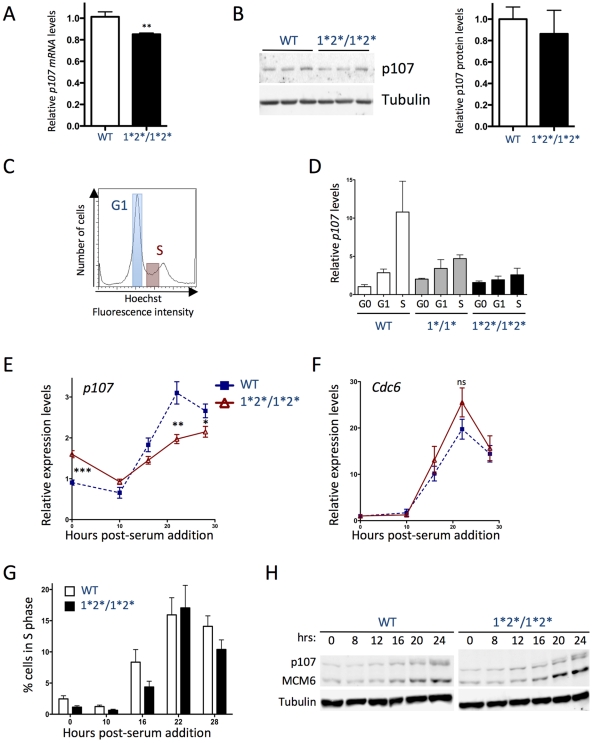
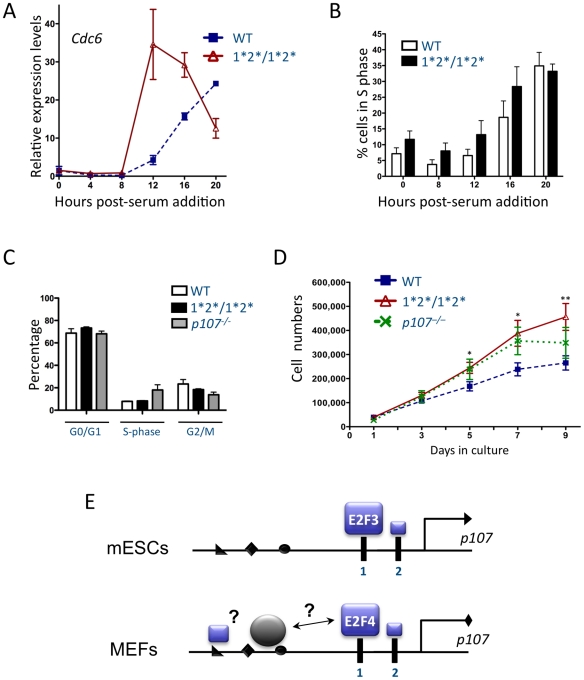
The retinoblastoma tumor suppressor (Rb) is a potent and ubiquitously expressed cell cycle regulator, but patients with a germline Rb mutation develop a very specific tumor spectrum. This surprising observation raises the possibility that mechanisms that compensate for loss of Rb function are present or activated in many cell types. In particular, p107, a protein related to Rb, has been shown to functionally overlap for loss of Rb in several cellular contexts. To investigate the mechanisms underlying this functional redundancy between Rb and p107 in vivo, we used gene targeting in embryonic stem cells to engineer point mutations in two consensus E2F binding sites in the endogenous p107 promoter. Analysis of normal and mutant cells by gene expression and chromatin immunoprecipitation assays showed that members of the Rb and E2F families directly bound these two sites. Furthermore, we found that these two E2F sites controlled both the repression of p107 in quiescent cells and also its activation in cycling cells, as well as in Rb mutant cells. Cell cycle assays further indicated that activation of p107 transcription during S phase through the two E2F binding sites was critical for controlled cell cycle progression, uncovering a specific role for p107 to slow proliferation in mammalian cells. Direct transcriptional repression of p107 by Rb and E2F family members provides a molecular mechanism for a critical negative feedback loop during cell cycle progression and tumorigenesis. These experiments also suggest novel therapeutic strategies to increase the p107 levels in tumor cells.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
E2F4-RB and E2F4-p107 complexes suppress gene expression by transforming growth factor beta through E2F binding sites.Proc Natl Acad Sci U S A. 1997 May 13;94(10):4948-53. doi: 10.1073/pnas.94.10.4948. Proc Natl Acad Sci U S A. 1997. PMID: 9144170 Free PMC article.
-
Retinoblastoma/p107/p130 pocket proteins: protein dynamics and interactions with target gene promoters.J Biol Chem. 2009 Jul 17;284(29):19265-71. doi: 10.1074/jbc.M808740200. Epub 2009 Mar 11. J Biol Chem. 2009. PMID: 19279001 Free PMC article.
-
The p107 tumor suppressor induces stable E2F DNA binding to repress target promoters.Oncogene. 2001 Apr 5;20(15):1882-91. doi: 10.1038/sj.onc.1204278. Oncogene. 2001. PMID: 11313936
-
From G0 to S phase: a view of the roles played by the retinoblastoma (Rb) family members in the Rb-E2F pathway.J Cell Biochem. 2007 Dec 15;102(6):1400-4. doi: 10.1002/jcb.21609. J Cell Biochem. 2007. PMID: 17979151 Review.
-
Regulation of the retinoblastoma-E2F pathway by the ubiquitin-proteasome system.Biochim Biophys Acta. 2015 Oct;1849(10):1289-97. doi: 10.1016/j.bbagrm.2015.08.008. Epub 2015 Aug 28. Biochim Biophys Acta. 2015. PMID: 26319102 Review.
Cited by
-
Retinoblastoma Protein Is Required for Epstein-Barr Virus Replication in Differentiated Epithelia.J Virol. 2023 Feb 28;97(2):e0103222. doi: 10.1128/jvi.01032-22. Epub 2023 Jan 31. J Virol. 2023. PMID: 36719239 Free PMC article.
-
Epstein-Barr virus replication within differentiated epithelia requires pRb sequestration of activator E2F transcription factors.J Virol. 2024 Oct 22;98(10):e0099524. doi: 10.1128/jvi.00995-24. Epub 2024 Sep 18. J Virol. 2024. PMID: 39291960 Free PMC article.
-
Critical role of the Rb family in myoblast survival and fusion.PLoS One. 2011 Mar 10;6(3):e17682. doi: 10.1371/journal.pone.0017682. PLoS One. 2011. PMID: 21423694 Free PMC article.
-
RBL1/p107 Expression Levels Are Modulated by Multiple Signaling Pathways.Cancers (Basel). 2021 Oct 8;13(19):5025. doi: 10.3390/cancers13195025. Cancers (Basel). 2021. PMID: 34638509 Free PMC article.
-
Essential role of CK2α for the interaction and stability of replication fork factors during DNA synthesis and activation of the S-phase checkpoint.Cell Mol Life Sci. 2022 Jun 4;79(6):339. doi: 10.1007/s00018-022-04374-3. Cell Mol Life Sci. 2022. PMID: 35661926 Free PMC article.
References
-
- Classon M, Dyson N. p107 and p130: versatile proteins with interesting pockets. Exp Cell Res. 2001;264:135–147. - PubMed
-
- Trimarchi JM, Lees JA. Sibling rivalry in the E2F family. Nat Rev Mol Cell Biol. 2002;3:11–20. - PubMed
-
- Ewen ME, Xing YG, Lawrence JB, Livingston DM. Molecular cloning, chromosomal mapping, and expression of the cDNA for p107, a retinoblastoma gene product-related protein. Cell. 1991;66:1155–1164. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
