

Identification of conserved regions and residues within Hedgehog acyltransferase critical for palmitoylation of Sonic Hedgehog
- PMID: 20585641
- PMCID: PMC2890405
- DOI: 10.1371/journal.pone.0011195
Identification of conserved regions and residues within Hedgehog acyltransferase critical for palmitoylation of Sonic Hedgehog
Abstract
Background: Sonic hedgehog (Shh) is a palmitoylated protein that plays key roles in mammalian development and human cancers. Palmitoylation of Shh is required for effective long and short range Shh-mediated signaling. Attachment of palmitate to Shh is catalyzed by Hedgehog acyltransferase (Hhat), a member of the membrane bound O-acyl transferase (MBOAT) family of multipass membrane proteins. The extremely hydrophobic composition of MBOAT proteins has limited their biochemical characterization. Except for mutagenesis of two conserved residues, there has been no structure-function analysis of Hhat, and the regions of the protein required for Shh palmitoylation are unknown.
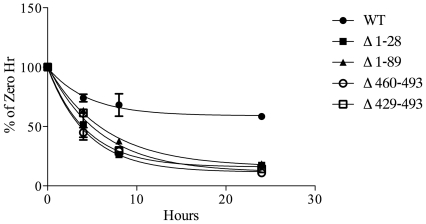
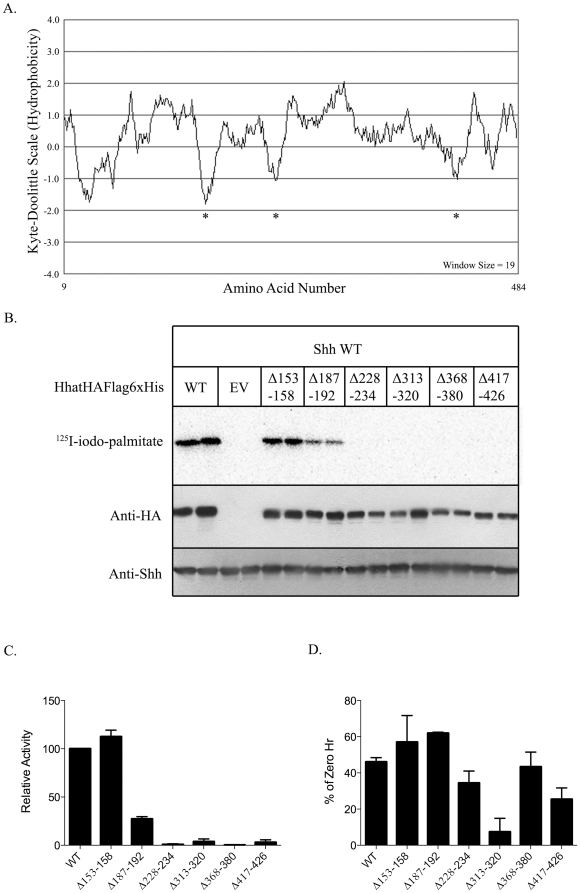
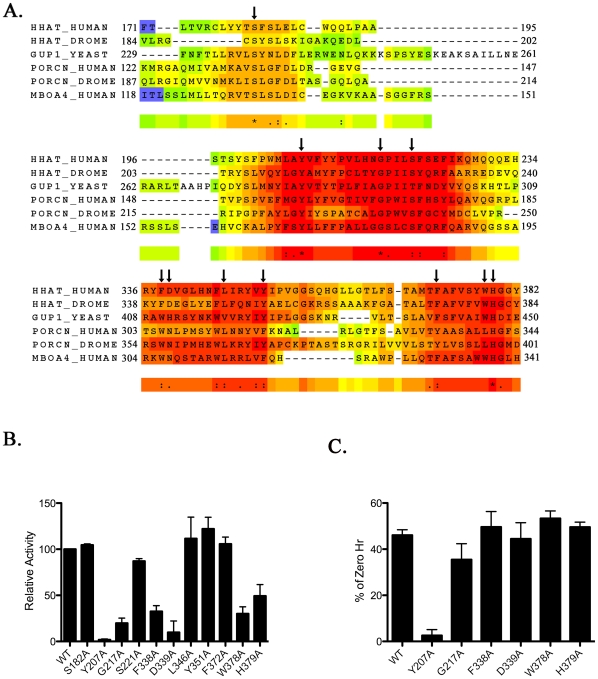
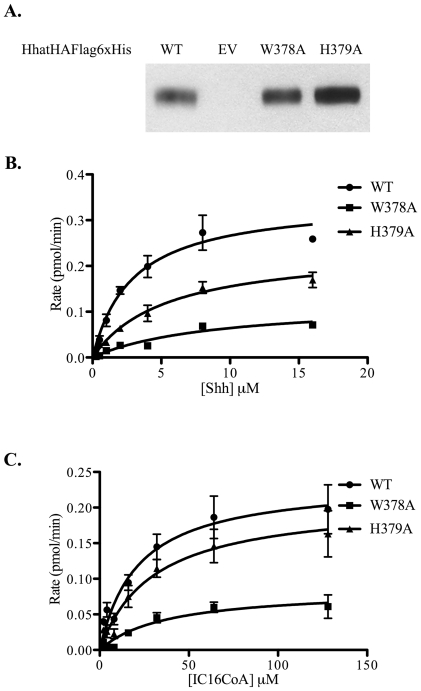
Methodology/principal findings: Here we undertake a systematic approach to identify residues within Hhat that are required for protein stability and/or enzymatic activity. We also identify a second, novel MBOAT homology region (residues 196-234) that is required for Hhat activity. In total, ten deletion mutants and eleven point mutants were generated and analyzed. Truncations at the N- and C-termini of Hhat yielded inactive proteins with reduced stability. Four Hhat mutants with deletions within predicted loop regions and five point mutants retained stability but lost palmitoylation activity. We purified two point mutants, W378A and H379A, with defective Hhat activity. Kinetic analyses revealed alterations in apparent K(m) and V(max) for Shh and/or palmitoyl CoA, changes that likely explain the catalytic defects observed for these mutants.
Conclusions/significance: This study has pinpointed specific regions and multiple residues that regulate Hhat stability and catalysis. Our findings should be applicable to other MBOAT proteins that mediate lipid modification of Wnt proteins and ghrelin, and should serve as a model for understanding how secreted morphogens are modified by palmitoyl acyltransferases.
Conflict of interest statement
Figures

References
-
- Ho KS, Scott MP. Sonic hedgehog in the nervous system: functions, modifications and mechanisms. Curr Opin Neurobiol. 2002;12:57–63. - PubMed
-
- Fuccillo M, Joyner AL, Fishell G. Morphogen to mitogen: the multiple roles of hedgehog signalling in vertebrate neural development. Nat Rev Neurosci. 2006;7:772–783. - PubMed
-
- McMahon AP, Ingham PW, Tabin CJ. Developmental roles and clinical significance of hedgehog signaling. Curr Top Dev Biol. 2003;53:1–114. - PubMed
-
- Lee JD, Kraus P, Gaiano N, Nery S, Kohtz J, et al. An acylatable residue of Hedgehog is differentially required in Drosophila and mouse limb development. Dev Biol. 2001;233:122–136. - PubMed
-
- Pasca di Magliano M, Hebrok M. Hedgehog signalling in cancer formation and maintenance. Nat Rev Cancer. 2003;3:903–911. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
