

Oriented single-crystal nuclear resonance vibrational spectroscopy of [Fe(TPP)(MI)(NO)]: quantitative assessment of the trans effect of NO
- PMID: 20586416
- PMCID: PMC2917100
- DOI: 10.1021/ic1010677
Oriented single-crystal nuclear resonance vibrational spectroscopy of [Fe(TPP)(MI)(NO)]: quantitative assessment of the trans effect of NO
Abstract
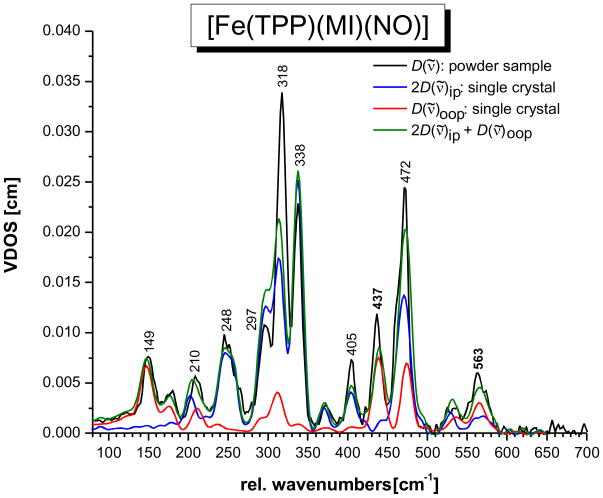
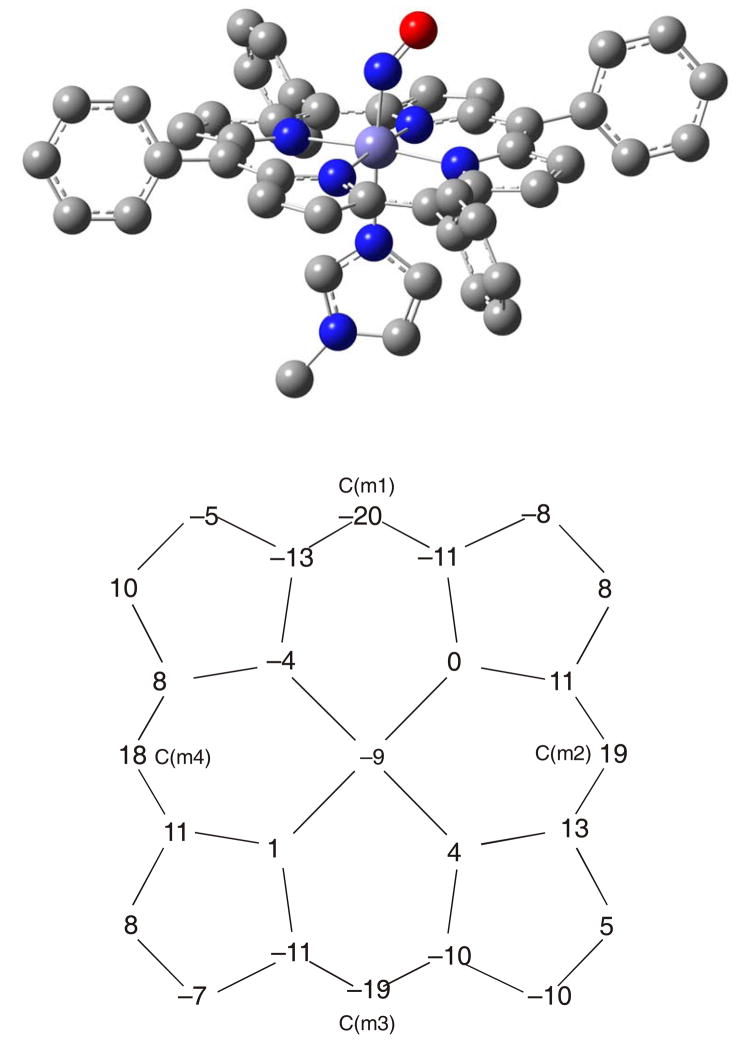
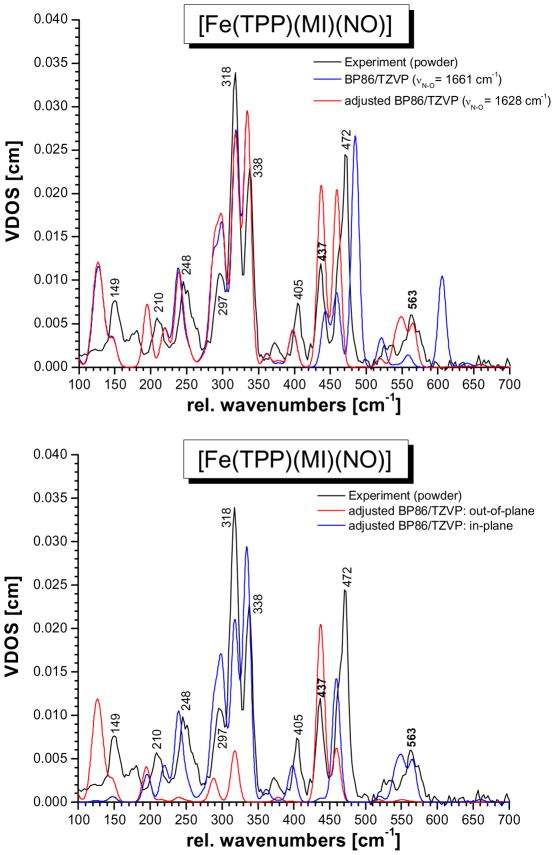
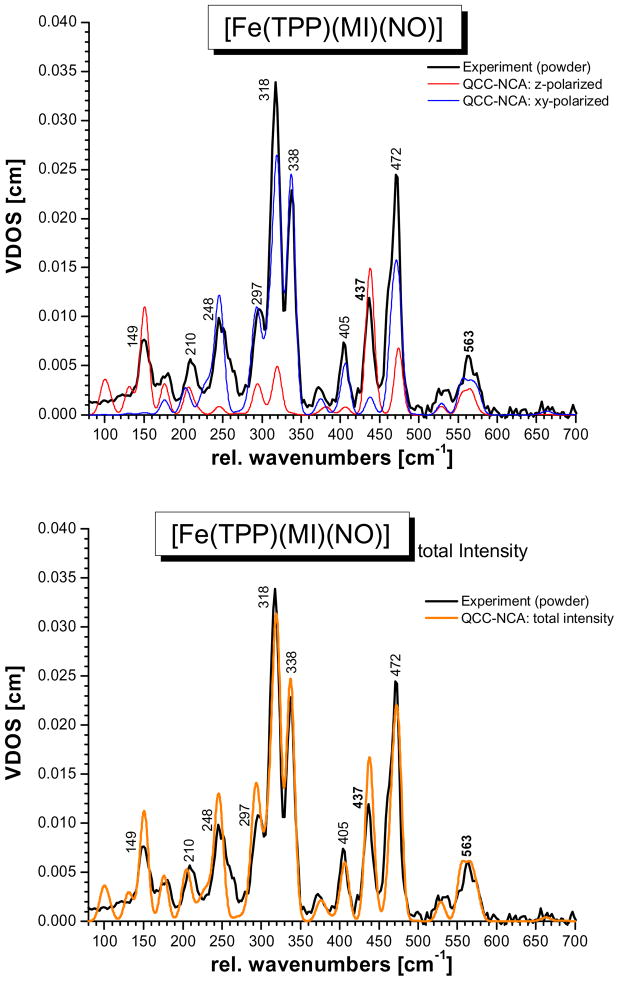
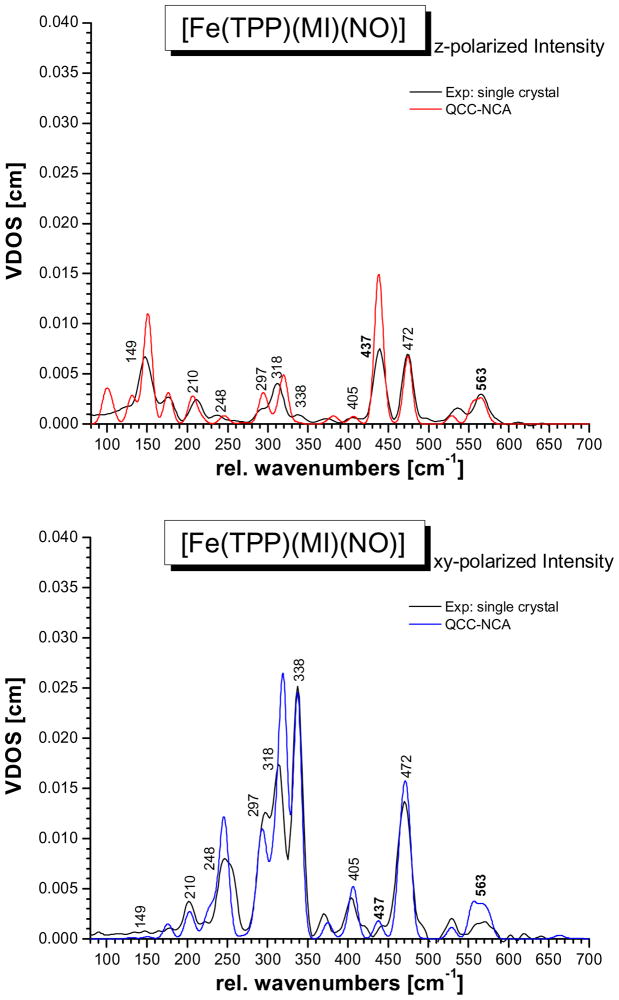
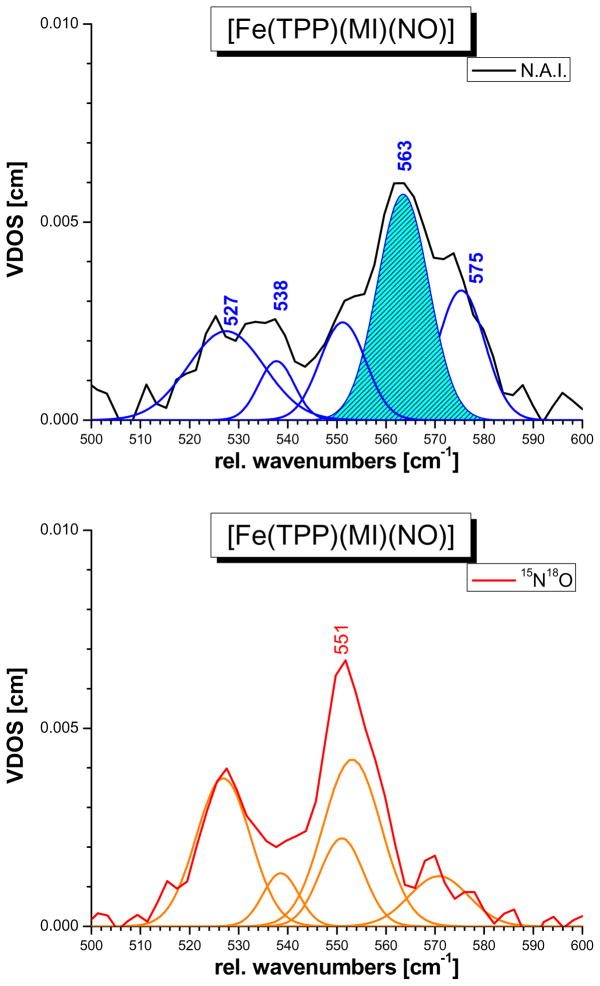
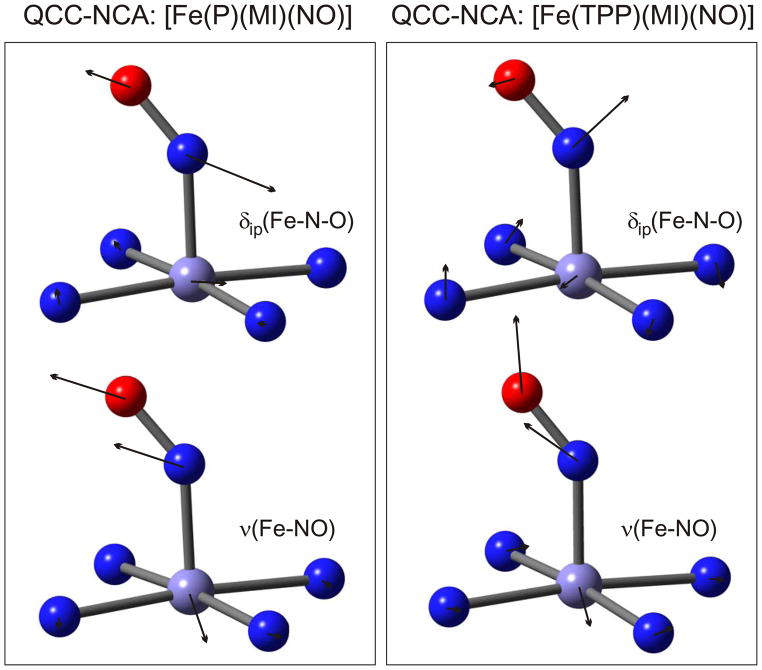
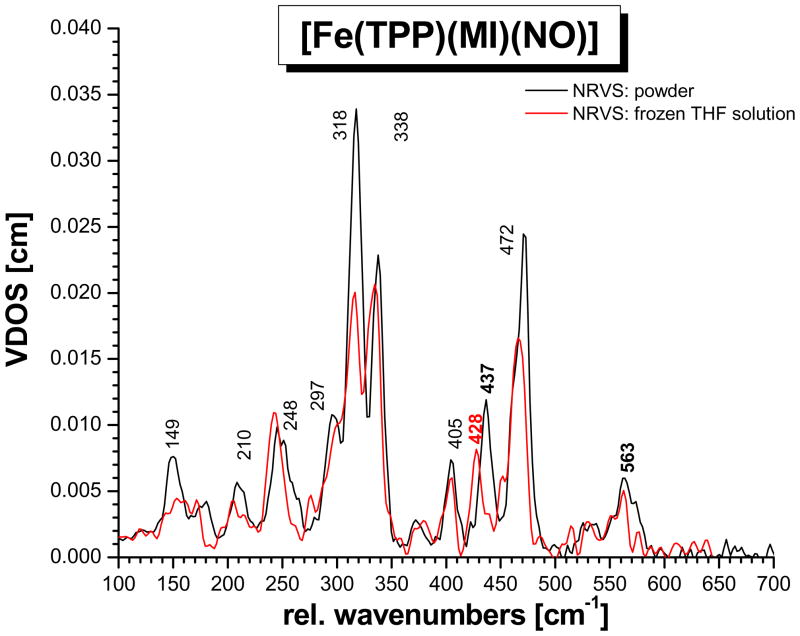
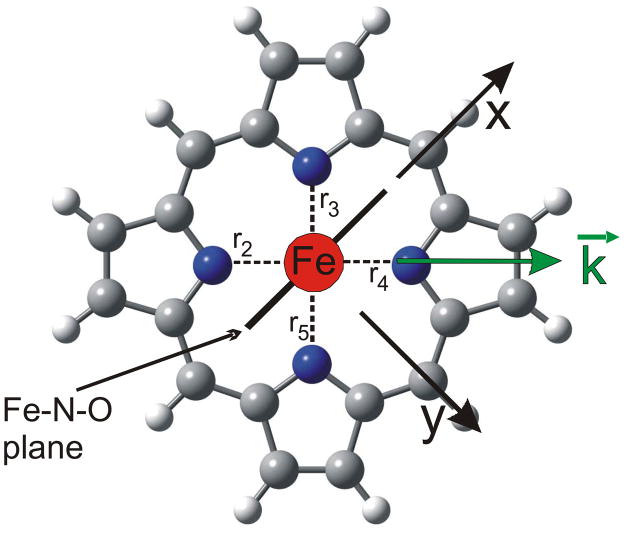
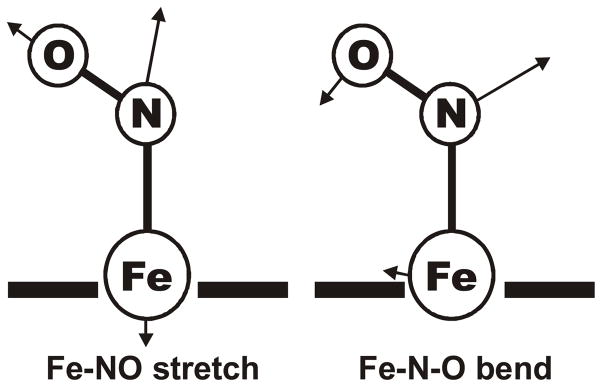
This paper presents oriented single-crystal Nuclear Resonance Vibrational Spectroscopy (NRVS) data for the six-coordinate (6C) ferrous heme-nitrosyl model complex [(57)Fe(TPP)(MI)(NO)] (1; TPP(2-) = tetraphenylporphyrin dianion; MI = 1-methylimidazole). The availability of these data enables for the first time the detailed simulation of the complete NRVS data, including the porphyrin-based vibrations, of a 6C ferrous heme-nitrosyl, using our quantum chemistry centered normal coordinate analysis (QCC-NCA). Importantly, the Fe-NO stretch is split by interaction with a porphyrin-based vibration into two features, observed at 437 and 472 cm(-1). The 437 cm(-1) feature is strongly out-of-plane (oop) polarized and shows a (15)N(18)O isotope shift of 8 cm(-1) and is therefore assigned to nu(Fe-NO). The admixture of Fe-N-O bending character is small. Main contributions to the Fe-N-O bend are observed in the 520-580 cm(-1) region, distributed over a number of in-plane (ip) polarized porphyrin-based vibrations. The main component, assigned to delta(ip)(Fe-N-O), is identified with the feature at 563 cm(-1). The Fe-N-O bend also shows strong mixing with the Fe-NO stretching internal coordinate, as evidenced by the oop NRVS intensity in the 520-580 cm(-1) region. Very accurate normal mode descriptions of nu(Fe-NO) and delta(ip)(Fe-N-O) have been obtained in this study. These results contradict previous interpretations of the vibrational spectra of 6C ferrous heme-nitrosyls where the higher energy feature at approximately 550 cm(-1) had usually been associated with nu(Fe-NO). Furthermore, these results provide key insight into NO binding to ferrous heme active sites in globins and other heme proteins, in particular with respect to (a) the effect of hydrogen bonding to the coordinated NO and (b) changes in heme dynamics upon NO coordination. [Fe(TPP)(MI)(NO)] constitutes an excellent model system for ferrous NO adducts of myoglobin (Mb) mutants where the distal histidine (His64) has been removed. Comparison to the reported vibrational data for wild-type (wt) Mb-NO then shows that the effect of H bonding to the coordinated NO is weak and mostly leads to a polarization of the pi/pi* orbitals of bound NO. In addition, the observation that delta(ip)(Fe-N-O) does not correlate well with nu(N-O) can be traced back to the very mixed nature of this mode. The Fe-N(imidazole) stretching frequency is observed at 149 cm(-1) in [Fe(TPP)(MI)(NO)], and spectral changes upon NO binding to five-coordinate ferrous heme active sites are discussed. The obtained high-quality force constants for the Fe-NO and N-O bonds of 2.57 and 11.55 mdyn/A can further be compared to those of corresponding 5C species, which allows for a quantitative analysis of the sigma trans interaction between the proximal imidazole (His) ligand and NO. This is key for the activation of the NO sensor soluble guanylate cyclase. Finally, DFT methods are calibrated against the experimentally determined vibrational properties of the Fe-N-O subunit in 1. DFT is in fact incapable of reproducing the vibrational energies and normal mode descriptions of the Fe-N-O unit well, and thus, DFT-based predictions of changes in vibrational properties upon heme modification or other perturbations of these 6C complexes have to be treated with caution.
Figures
Similar articles
-
Nuclear resonance vibrational spectroscopy applied to [Fe(OEP)(NO)]: the vibrational assignments of five-coordinate ferrous heme-nitrosyls and implications for electronic structure.Inorg Chem. 2010 May 3;49(9):4133-48. doi: 10.1021/ic902181e. Inorg Chem. 2010. PMID: 20345089 Free PMC article.
-
Axial ligand effects on vibrational dynamics of iron in heme carbonyl studied by nuclear resonance vibrational spectroscopy.J Phys Chem B. 2012 Nov 29;116(47):13831-8. doi: 10.1021/jp304398g. Epub 2012 Nov 14. J Phys Chem B. 2012. PMID: 23072485
-
Electronic structure of six-coordinate iron(III)-porphyrin NO adducts: the elusive iron(III)-NO(radical) state and its influence on the properties of these complexes.J Am Chem Soc. 2008 Nov 19;130(46):15288-303. doi: 10.1021/ja801860u. Epub 2008 Oct 23. J Am Chem Soc. 2008. PMID: 18942830
-
Electronic structure of heme-nitrosyls and its significance for nitric oxide reactivity, sensing, transport, and toxicity in biological systems.Inorg Chem. 2010 Jul 19;49(14):6293-316. doi: 10.1021/ic902304a. Inorg Chem. 2010. PMID: 20666388 Review.
-
Nuclear resonance vibrational spectroscopy--NRVS.J Inorg Biochem. 2005 Jan;99(1):60-71. doi: 10.1016/j.jinorgbio.2004.11.004. J Inorg Biochem. 2005. PMID: 15598492 Review.
Cited by
-
Anisotropic iron motion in nitrosyl iron porphyrinates: natural and synthetic hemes.Inorg Chem. 2014 Mar 3;53(5):2582-90. doi: 10.1021/ic4028964. Epub 2014 Feb 14. Inorg Chem. 2014. PMID: 24528178 Free PMC article.
-
The diagnostic vibrational signature of pentacoordination in heme carbonyls.J Am Chem Soc. 2014 Jul 16;136(28):9818-21. doi: 10.1021/ja503191z. Epub 2014 Jul 7. J Am Chem Soc. 2014. PMID: 24950373 Free PMC article.
-
Nuclear inelastic scattering and Mössbauer spectroscopy as local probes for ligand binding modes and electronic properties in proteins: vibrational behavior of a ferriheme center inside a β-barrel protein.J Am Chem Soc. 2012 Mar 7;134(9):4216-28. doi: 10.1021/ja210067t. Epub 2012 Feb 27. J Am Chem Soc. 2012. PMID: 22295945 Free PMC article.
-
3D Motions of Iron in Six-Coordinate {FeNO}(7) Hemes by Nuclear Resonance Vibration Spectroscopy.Chemistry. 2016 Apr 25;22(18):6323-6332. doi: 10.1002/chem.201505155. Epub 2016 Mar 21. Chemistry. 2016. PMID: 26999733 Free PMC article.
-
SciPhon: a data analysis software for nuclear resonant inelastic X-ray scattering with applications to Fe, Kr, Sn, Eu and Dy.J Synchrotron Radiat. 2018 Sep 1;25(Pt 5):1581-1599. doi: 10.1107/S1600577518009487. Epub 2018 Aug 21. J Synchrotron Radiat. 2018. PMID: 30179200 Free PMC article.
References
-
- Moncada S, Palmer RM, Higgs EA. Pharmacol Rev. 1991;43:109–142. - PubMed
- Snyder SH. Science. 1992;257:494–496. - PubMed
- Butler AR, Williams DLH. Chem Soc Revs. 1993:233–241.
- Bredt DS, Snyder SH. Annu Rev Biochem. 1994;63:175–195. - PubMed
- Lancaster JR., Jr . In: Encyclopedia of Inorganic Chemistry. Bruce RB, editor. Wiley; Chichester: 1994.
- Feelisch M, Stamler JS, editors. Methods in Nitric Oxide Research. Wiley; Chichester: 1996.
- Ignarro L, editor. Nitric Oxide: Biology and Pathobiology. Academic Press; San Diego: 2000.
-
- Stuehr DJ. Annu Rev Pharmacol Toxicol. 1997;37:339–359. - PubMed
- Raman CS, Li H, Martasek P, Kral V, Masters BSS, Poulos TL. Cell. 1998;95:939–950. - PubMed
- Fischmann TO, Hruza A, Niu XD, Fossetta JD, Lunn CA, Dolphin E, Prongay AJ, Reichert P, Lundell DJ, Narula SK, Weber PC. Nat Struct Biol. 1999;6:233–242. - PubMed
- Li H, Poulos TL. J Inorg Biochem. 2005;99:293–305. - PubMed
- Rousseau DL, Li D, Couture M, Yeh SR. J Inorg Biochem. 2005;99:306–323. - PubMed
- Martin NI, Woodward JJ, Winter MB, Beeson WT, Marletta MA. J Am Chem Soc. 2007;129:12563–12570. - PubMed
-
- Garbers DL, Lowe DG. J Biol Chem. 1994;269:30741–30744. - PubMed
- Zhao Y, Hoganson C, Babcock GT, Marletta MA. Biochemistry. 1998;37:12458–12464. - PubMed
- Zhao Y, Brandish PE, Ballou DP, Marletta MA. Proc Natl Acad Sci USA. 1999;96:14753–14758. - PMC - PubMed
- Ballou DP, Zhao Y, Brandish PE, Marletta MA. Proc Natl Acad Sci USA. 2002;99:12097–12101. - PMC - PubMed
- Karow DS, Pan D, Tran R, Pellicena P, Presley A, Mathies RA, Marletta MA. Biochemistry. 2004;43:10203–10211. - PubMed
- Boon EM, Marletta MA. Curr Opin Chem Biol. 2005;9:441–446. - PubMed
- Gilles-Gonzalez MA, Gonzales G. J Inorg Biochem. 2005;99:1–22. - PubMed
-
- Cary SPL, Winger JA, Derbyshire ER, Marletta MA. TRENDS Biochem Sci. 2006;31:231–239. - PubMed
-
- Scheidt WR, Piciulo PL. J Am Chem Soc. 1976;98:1913–1919. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
