

Comparative analysis of copy number detection by whole-genome BAC and oligonucleotide array CGH
- PMID: 20587050
- PMCID: PMC2909945
- DOI: 10.1186/1755-8166-3-11
Comparative analysis of copy number detection by whole-genome BAC and oligonucleotide array CGH
Abstract
Background: Microarray-based comparative genomic hybridization (aCGH) is a powerful diagnostic tool for the detection of DNA copy number gains and losses associated with chromosome abnormalities, many of which are below the resolution of conventional chromosome analysis. It has been presumed that whole-genome oligonucleotide (oligo) arrays identify more clinically significant copy-number abnormalities than whole-genome bacterial artificial chromosome (BAC) arrays, yet this has not been systematically studied in a clinical diagnostic setting.
Results: To determine the difference in detection rate between similarly designed BAC and oligo arrays, we developed whole-genome BAC and oligonucleotide microarrays and validated them in a side-by-side comparison of 466 consecutive clinical specimens submitted to our laboratory for aCGH. Of the 466 cases studied, 67 (14.3%) had a copy-number imbalance of potential clinical significance detectable by the whole-genome BAC array, and 73 (15.6%) had a copy-number imbalance of potential clinical significance detectable by the whole-genome oligo array. However, because both platforms identified copy number variants of unclear clinical significance, we designed a systematic method for the interpretation of copy number alterations and tested an additional 3,443 cases by BAC array and 3,096 cases by oligo array. Of those cases tested on the BAC array, 17.6% were found to have a copy-number abnormality of potential clinical significance, whereas the detection rate increased to 22.5% for the cases tested by oligo array. In addition, we validated the oligo array for detection of mosaicism and found that it could routinely detect mosaicism at levels of 30% and greater.
Conclusions: Although BAC arrays have faster turnaround times, the increased detection rate of oligo arrays makes them attractive for clinical cytogenetic testing.
Figures

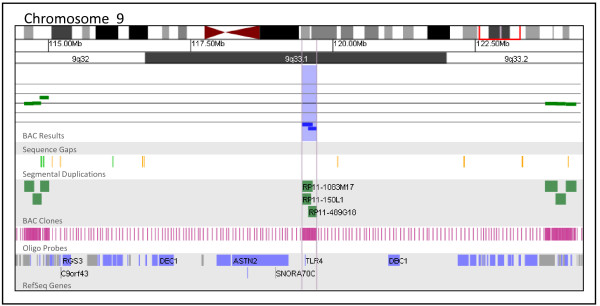
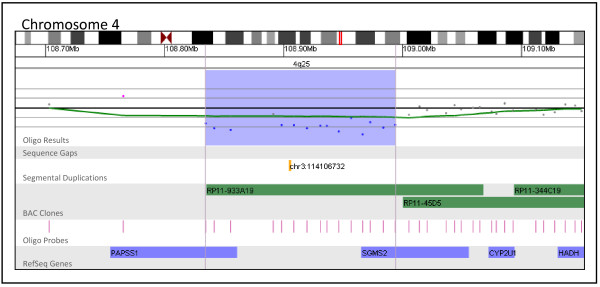
References
-
- Shaffer LG, Bejjani BA, Torchia B, Kirkpatrick S, Coppinger J, Ballif BC. The identification of microdeletion syndromes and other chromosome abnormalities: Cytogenetic methods of the past, new technologies for the future. Am J Med Genet C Semin Med Genet. 2007;145C:335–345. doi: 10.1002/ajmg.c.30152. - DOI - PubMed
-
- Ballif BC, Hornor SA, Jenkins E, Madan-Khetarpal S, Surti U, Jackson KE, Asamoah A, Brock PL, Gowans GC, Conway RL, Graham JM Jr, Medne L, Zackai EH, Shaikh TH, Geoghegan J, Selzer RR, Eis PS, Bejjani BA, Shaffer LG. Discovery of a previously unrecognized microdeletion syndrome of 16p11.2-p12.2. Nat Genet. 2007;39:1071–1073. doi: 10.1038/ng2107. - DOI - PubMed
-
- Koolen DA, Vissers LE, Pfundt R, de Leeuw N, Knight SJ, Regan R, Kooy RF, Reyniers E, Romano C, Fichera M, Schinzel A, Baumer A, Anderlid BM, Schoumans J, Knoers NV, van Kessel AG, Sistermans EA, Veltman JA, Brunner HG, de Vries BB. A new chromosome 17q21.31 microdeletion syndrome associated with a common inversion polymorphism. Nat Genet. 2006;38:999–1001. doi: 10.1038/ng1853. - DOI - PubMed
LinkOut - more resources
Full Text Sources
