

Susceptible cell lines for the production of porcine reproductive and respiratory syndrome virus by stable transfection of sialoadhesin and CD163
- PMID: 20587060
- PMCID: PMC2908558
- DOI: 10.1186/1472-6750-10-48
Susceptible cell lines for the production of porcine reproductive and respiratory syndrome virus by stable transfection of sialoadhesin and CD163
Abstract
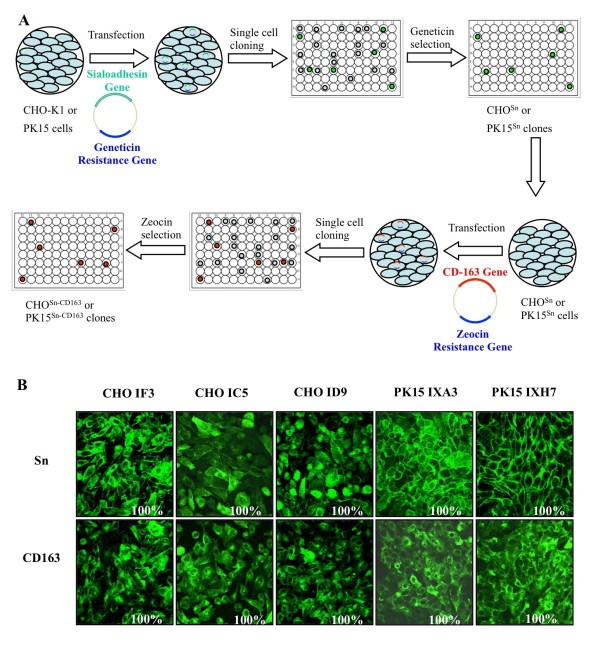
Background: Porcine reproductive and respiratory syndrome virus (PRRSV) causes major economic losses in the pig industry worldwide. In vivo, the virus infects a subpopulation of tissue macrophages. In vitro, PRRSV only replicates in primary pig macrophages and African green monkey kidney derived cells, such as Marc-145. The latter is currently used for vaccine production. However, since virus entry in Marc-145 cells is different compared to entry in primary macrophages, specific epitopes associated with virus entry could potentially alter upon growth on Marc-145 cells. To avoid this, we constructed CHO and PK15 cell lines recombinantly expressing the PRRSV receptors involved in virus entry into macrophages, sialoadhesin (Sn) and CD163 (CHOSn-CD163 and PK15Sn-CD163) and evaluated their potential for production of PRRSV.
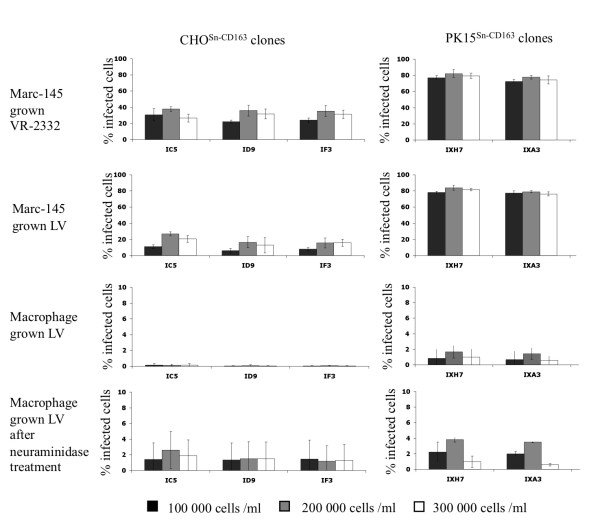
Results: Detailed analysis of PRRSV infection revealed that LV and VR-2332 virus particles could attach to and internalize into the CHOSn-CD163 and PK15Sn-CD163 cells. Initially, this occurred less efficiently for macrophage grown virus than for Marc-145 grown virus. Upon internalization, disassembly of the virus particles was observed. The two cell lines could be infected with PRRSV strains LV and VR-2332. However, it was observed that Marc-145 grown virus infected the cells more efficiently than macrophage grown virus. If the cells were treated with neuraminidase to remove cis-acting sialic acids that hinder the interaction of the virus with Sn, the amount of infected cells with macrophage grown virus increased. Comparison of both cell lines showed that the PK15Sn-CD163 cell line gave in general better results than the CHOSn-CD163 cell line. Only 2 out of 5 PRRSV strains replicated well in CHOSn-CD163 cells. Furthermore, the virus titer of all 5 PRRSV strains produced after passaging in PK15Sn-CD163 cells was similar to the virus titer of those strains produced in Marc-145 cells. Analysis of the sequence of the structural proteins of original virus and virus grown for 5 passages on PK15Sn-CD163 cells showed either no amino acid (aa) changes (VR-2332 and 07V063), one aa (LV), two aa (08V194) or three aa (08V204) changes. None of these changes are situated in known neutralizing epitopes.
Conclusions: A PRRSV susceptible cell line was constructed that can grow virus to similar levels compared to currently available cell lines. Mutations induced by growth on this cell lines were either absent or minimal and located outside known neutralizing epitopes. Together, the results show that this cell line can be used to produce vaccine virus and for PRRSV virus isolation.
Figures
Similar articles
-
Comparison of Primary Virus Isolation in Pulmonary Alveolar Macrophages and Four Different Continuous Cell Lines for Type 1 and Type 2 Porcine Reproductive and Respiratory Syndrome Virus.Vaccines (Basel). 2021 Jun 3;9(6):594. doi: 10.3390/vaccines9060594. Vaccines (Basel). 2021. PMID: 34205087 Free PMC article.
-
Sialoadhesin and CD163 join forces during entry of the porcine reproductive and respiratory syndrome virus.J Gen Virol. 2008 Dec;89(Pt 12):2943-2953. doi: 10.1099/vir.0.2008/005009-0. J Gen Virol. 2008. PMID: 19008379
-
CD163 expression confers susceptibility to porcine reproductive and respiratory syndrome viruses.J Virol. 2007 Jul;81(14):7371-9. doi: 10.1128/JVI.00513-07. Epub 2007 May 9. J Virol. 2007. PMID: 17494075 Free PMC article.
-
A brief review of CD163 and its role in PRRSV infection.Virus Res. 2010 Dec;154(1-2):98-103. doi: 10.1016/j.virusres.2010.07.018. Epub 2010 Jul 23. Virus Res. 2010. PMID: 20655964 Review.
-
PRRS virus receptors and their role for pathogenesis.Vet Microbiol. 2015 Jun 12;177(3-4):229-41. doi: 10.1016/j.vetmic.2015.04.002. Epub 2015 Apr 13. Vet Microbiol. 2015. PMID: 25912022 Review.
Cited by
-
Identification of New Compounds against PRRSV Infection by Directly Targeting CD163.J Virol. 2023 May 31;97(5):e0005423. doi: 10.1128/jvi.00054-23. Epub 2023 May 3. J Virol. 2023. PMID: 37133376 Free PMC article.
-
Comparison of Primary Virus Isolation in Pulmonary Alveolar Macrophages and Four Different Continuous Cell Lines for Type 1 and Type 2 Porcine Reproductive and Respiratory Syndrome Virus.Vaccines (Basel). 2021 Jun 3;9(6):594. doi: 10.3390/vaccines9060594. Vaccines (Basel). 2021. PMID: 34205087 Free PMC article.
-
Developing a Triple Transgenic Cell Line for High-Efficiency Porcine Reproductive and Respiratory Syndrome Virus Infection.PLoS One. 2016 May 16;11(5):e0154238. doi: 10.1371/journal.pone.0154238. eCollection 2016. PLoS One. 2016. PMID: 27182980 Free PMC article.
-
No Evidence for a Role for Antibodies during Vaccination-Induced Enhancement of Porcine Reproductive and Respiratory Syndrome.Viruses. 2019 Sep 6;11(9):829. doi: 10.3390/v11090829. Viruses. 2019. PMID: 31489915 Free PMC article.
-
Differential innate immune response of endometrial cells to porcine reproductive and respiratory syndrome virus type 1 versus type 2.PLoS One. 2023 Apr 26;18(4):e0284658. doi: 10.1371/journal.pone.0284658. eCollection 2023. PLoS One. 2023. PMID: 37099541 Free PMC article.
References
-
- Cavanagh D. Nidovirales: a new order comprising Coronaviridae and Arteriviridae. Archives of virology. 1997;142(3):629–633. - PubMed
-
- Snijder EJ, Meulenberg JJ. The molecular biology of arteriviruses. The Journal of general virology. 1998;79(Pt 5):961–979. - PubMed
-
- Neumann EJ, Kliebenstein JB, Johnson CD, Mabry JW, Bush EJ, Seitzinger AH, Green AL, Zimmerman JJ. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. Journal of the American Veterinary Medical Association. 2005;227(3):385–392. doi: 10.2460/javma.2005.227.385. - DOI - PubMed
-
- Christianson WT, Choi CS, Collins JE, Molitor TW, Morrison RB, Joo HS. Pathogenesis of porcine reproductive and respiratory syndrome virus infection in mid-gestation sows and fetuses. Canadian journal of veterinary research = Revue canadienne de recherche veterinaire. 1993;57(4):262–268. - PMC - PubMed
-
- Christianson WT, Collins JE, Benfield DA, Harris L, Gorcyca DE, Chladek DW, Morrison RB, Joo HS. Experimental reproduction of swine infertility and respiratory syndrome in pregnant sows. American journal of veterinary research. 1992;53(4):485–488. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
