

Confocal Analysis of Cholinergic and Dopaminergic Inputs onto Pyramidal Cells in the Prefrontal Cortex of Rodents
- PMID: 20589096
- PMCID: PMC2893003
- DOI: 10.3389/fnana.2010.00021
Confocal Analysis of Cholinergic and Dopaminergic Inputs onto Pyramidal Cells in the Prefrontal Cortex of Rodents
Abstract
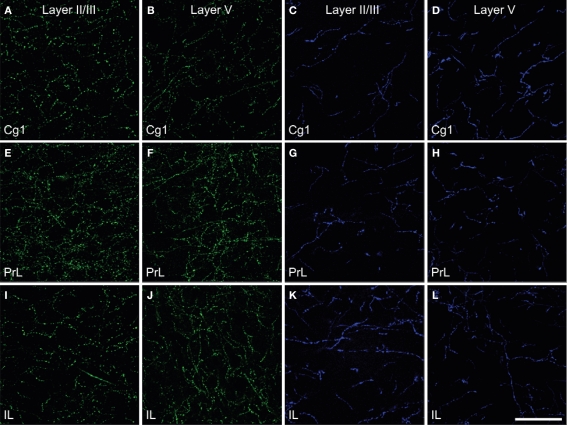
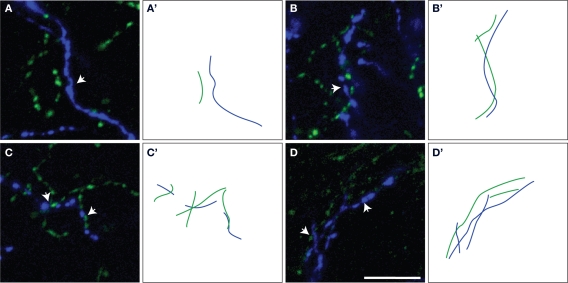
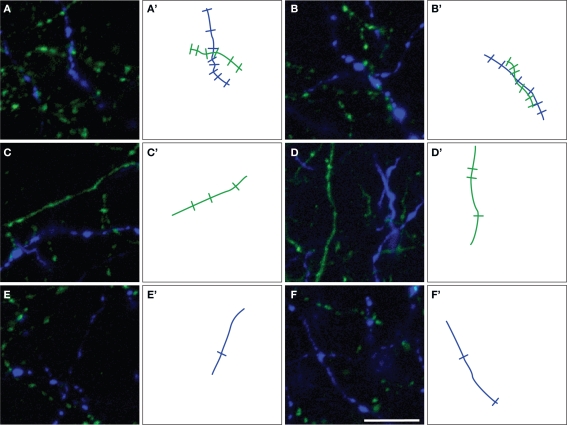
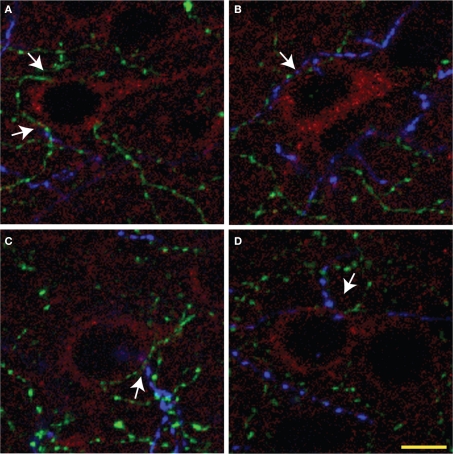
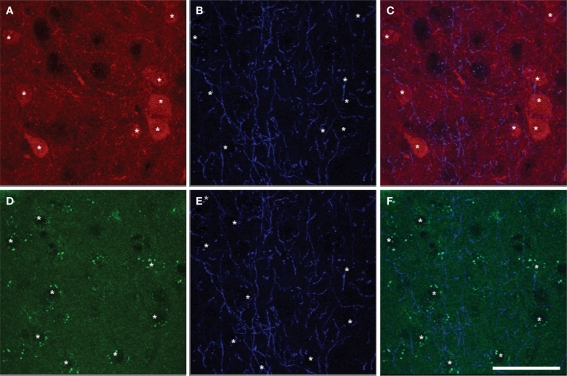
Cholinergic and dopaminergic projections to the rat medial prefrontal cortex (mPFC) are both involved in cognitive functions including attention. These neuronal systems modulate mPFC neuronal activity mainly through diffuse transmission. In order to better understand the anatomical level of influence of these systems, confocal microscopy with triple-fluorescent immunolabeling was used in three subregions of the mPFC of rats and Drd1a-tdTomato/Drd2-EGFP transgenic mice. The zone of interaction was defined as a reciprocal microproximity between dopaminergic and cholinergic axonal segments as well as pyramidal neurons. The density of varicosities, along these segments was considered as a possible activity-dependant morphological feature. The percentage of cholinergic and dopaminergic fibers in microproximity ranged from 12 to 40% depending on the layer and mPFC subregion. The cholinergic system appeared to have more influence on dopaminergic fibers since a larger proportion of the dopaminergic fibers were within microproximity to cholinergic fibers. The density of both cholinergic and dopaminergic varicosities was significantly elevated within microproximities. The main results indicate that the cholinergic and dopaminergic systems converge on pyramidal cells in mPFC particularly in the layer V. In transgenic mice 93% of the pyramidal cells expressed the transgenic marker for Drd2 expression, but only 22% expressed the maker for Drd1ar expression. Data presented here suggest that the modulation of mPFC by dopaminergic fibers would be mostly inhibitory and localized at the output level whereas the cholinergic modulation would be exerted at the input and output level both through direct interaction with pyramidal cells and dopaminergic fibers.
Keywords: acetylcholine; cingulate cortex; diffuse transmission; dopamine; infralimbic cortex; prelimbic cortex.
Figures
Similar articles
-
Axonal varicosity density as an index of local neuronal interactions.PLoS One. 2011;6(7):e22543. doi: 10.1371/journal.pone.0022543. Epub 2011 Jul 21. PLoS One. 2011. PMID: 21811630 Free PMC article.
-
Activation of the mouse primary visual cortex by medial prefrontal subregion stimulation is not mediated by cholinergic basalo-cortical projections.Front Syst Neurosci. 2015 Feb 9;9:1. doi: 10.3389/fnsys.2015.00001. eCollection 2015. Front Syst Neurosci. 2015. PMID: 25709570 Free PMC article.
-
Subregion preference in the long-range connectome of pyramidal neurons in the medial prefrontal cortex.BMC Biol. 2024 Apr 29;22(1):95. doi: 10.1186/s12915-024-01880-7. BMC Biol. 2024. PMID: 38679719 Free PMC article.
-
Neurotransmitter systems of the medial prefrontal cortex: potential role in sensitization to psychostimulants.Brain Res Brain Res Rev. 2003 Mar;41(2-3):203-28. doi: 10.1016/s0165-0173(02)00233-3. Brain Res Brain Res Rev. 2003. PMID: 12663081 Review.
-
Dopaminergic modulation of pain signals in the medial prefrontal cortex: Challenges and perspectives.Neurosci Lett. 2019 May 29;702:71-76. doi: 10.1016/j.neulet.2018.11.043. Epub 2018 Nov 29. Neurosci Lett. 2019. PMID: 30503912 Review.
Cited by
-
The Development of the Mesoprefrontal Dopaminergic System in Health and Disease.Front Neural Circuits. 2021 Oct 12;15:746582. doi: 10.3389/fncir.2021.746582. eCollection 2021. Front Neural Circuits. 2021. PMID: 34712123 Free PMC article. Review.
-
Disentangling the diverse roles of dopamine D2 receptors in striatal function and behavior.Neurochem Int. 2019 May;125:35-46. doi: 10.1016/j.neuint.2019.01.022. Epub 2019 Feb 1. Neurochem Int. 2019. PMID: 30716356 Free PMC article. Review.
-
Noradrenalin and dopamine receptors both control cAMP-PKA signaling throughout the cerebral cortex.Front Cell Neurosci. 2014 Aug 21;8:247. doi: 10.3389/fncel.2014.00247. eCollection 2014. Front Cell Neurosci. 2014. PMID: 25191229 Free PMC article.
-
Impaired Functional Connectivity in the Prefrontal Cortex: A Mechanism for Chronic Stress-Induced Neuropsychiatric Disorders.Neural Plast. 2016;2016:7539065. doi: 10.1155/2016/7539065. Epub 2016 Jan 19. Neural Plast. 2016. PMID: 26904302 Free PMC article. Review.
-
The Association between α-Synuclein and α-Tubulin in Brain Synapses.Int J Mol Sci. 2021 Aug 25;22(17):9153. doi: 10.3390/ijms22179153. Int J Mol Sci. 2021. PMID: 34502063 Free PMC article.
References
-
- Broersen L. M., Heinsbroek R. P., de Bruin J. P., Uylings H. B., Olivier B. (1995). The role of the medial prefrontal cortex of rats in short-term memory functioning: further support for involvement of cholinergic, rather than dopaminergic mechanisms. Brain Res. 674, 221–22910.1016/0006-8993(95)00025-L - DOI - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases