

Nonintegrating foamy virus vectors
- PMID: 20592072
- PMCID: PMC2937610
- DOI: 10.1128/JVI.00394-10
Nonintegrating foamy virus vectors
Abstract
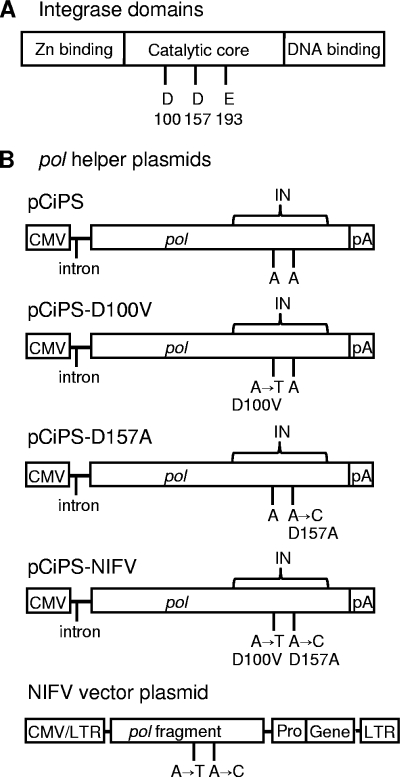
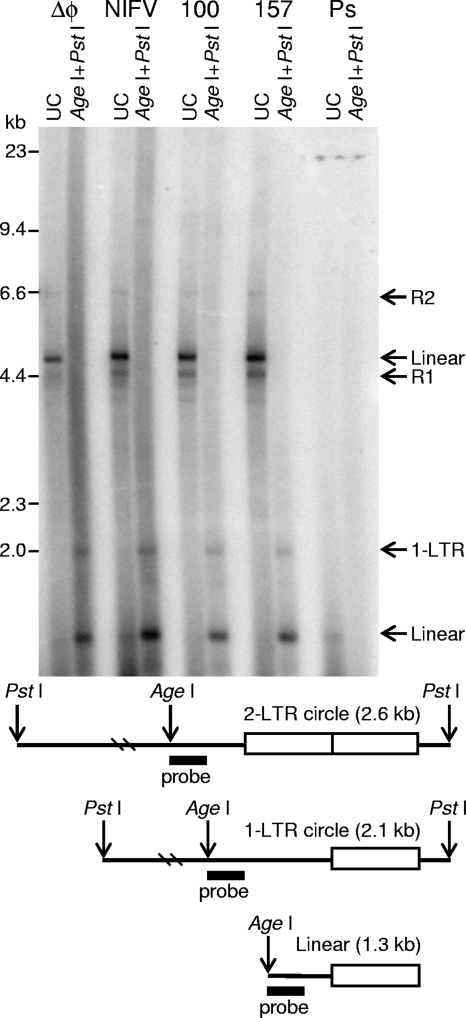
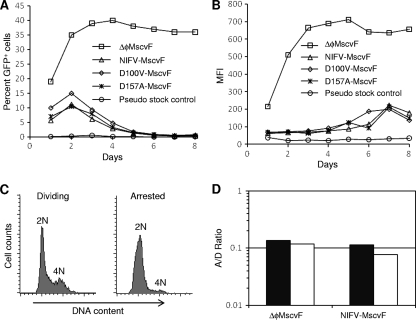
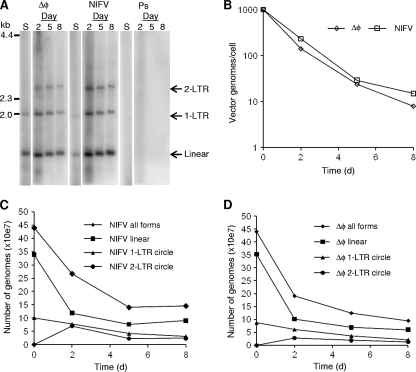
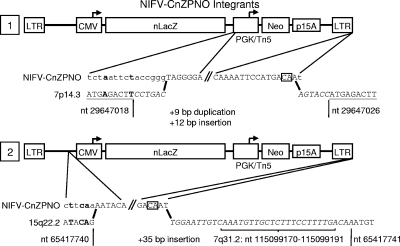
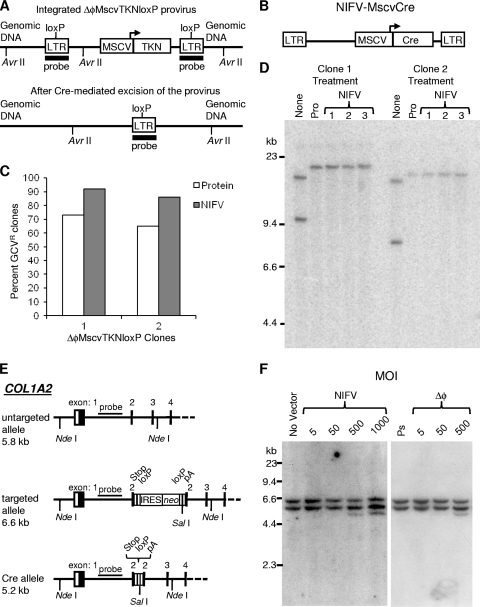
Foamy viruses (FVs), or spumaviruses, are integrating retroviruses that have been developed as vectors. Here we generated nonintegrating foamy virus (NIFV) vectors by introducing point mutations into the highly conserved DD35E catalytic core motif of the foamy virus integrase sequence. NIFV vectors produced high-titer stocks, transduced dividing cells, and did not integrate. Cells infected with NIFV vectors contained episomal vector genomes that consisted of linear, 1-long-terminal-repeat (1-LTR), and 2-LTR circular DNAs. These episomes expressed transgenes, were stable, and became progressively diluted in the dividing cell population. 1-LTR circles but not 2-LTR circles were found in all vector stocks prior to infection. Residual integration of NIFV vectors occurred at a frequency 4 logs lower than that of integrase-proficient FV vectors. Cre recombinase expressed from a NIFV vector mediated excision of both an integrated, floxed FV vector and a gene-targeted neo expression cassette, demonstrating the utility of these episomal vectors. The broad host range and large packaging capacity of NIFV vectors should make them useful for a variety of applications requiring transient gene expression.
Figures
Similar articles
-
Foamy Virus Integrase in Development of Viral Vector for Gene Therapy.J Microbiol Biotechnol. 2020 Sep 28;30(9):1273-1281. doi: 10.4014/jmb.2003.03046. J Microbiol Biotechnol. 2020. PMID: 32699199 Free PMC article. Review.
-
Production of foamy virus vector and transduction of hematopoietic cells.Cold Spring Harb Protoc. 2010 Sep 1;2010(9):pdb.prot5481. doi: 10.1101/pdb.prot5481. Cold Spring Harb Protoc. 2010. PMID: 20810629 Free PMC article.
-
Foamy Virus Vector Carries a Strong Insulator in Its Long Terminal Repeat Which Reduces Its Genotoxic Potential.J Virol. 2017 Dec 14;92(1):e01639-17. doi: 10.1128/JVI.01639-17. Print 2018 Jan 1. J Virol. 2017. PMID: 29046446 Free PMC article.
-
A novel function for spumaretrovirus integrase: an early requirement for integrase-mediated cleavage of 2 LTR circles.Retrovirology. 2005 May 18;2:31. doi: 10.1186/1742-4690-2-31. Retrovirology. 2005. PMID: 15904533 Free PMC article.
-
Advances in foamy virus vector systems: Development and applications.Virology. 2025 Jan;601:110270. doi: 10.1016/j.virol.2024.110270. Epub 2024 Oct 23. Virology. 2025. PMID: 39509861 Review.
Cited by
-
Histone deacetylase inhibition activates transgene expression from integration-defective lentiviral vectors in dividing and non-dividing cells.Hum Gene Ther. 2013 Jan;24(1):78-96. doi: 10.1089/hum.2012.069. Epub 2012 Dec 11. Hum Gene Ther. 2013. PMID: 23140481 Free PMC article.
-
Foamy Virus Integrase in Development of Viral Vector for Gene Therapy.J Microbiol Biotechnol. 2020 Sep 28;30(9):1273-1281. doi: 10.4014/jmb.2003.03046. J Microbiol Biotechnol. 2020. PMID: 32699199 Free PMC article. Review.
-
Normal collagen and bone production by gene-targeted human osteogenesis imperfecta iPSCs.Mol Ther. 2012 Jan;20(1):204-13. doi: 10.1038/mt.2011.209. Epub 2011 Oct 25. Mol Ther. 2012. PMID: 22031238 Free PMC article.
-
Silent IL2RG Gene Editing in Human Pluripotent Stem Cells.Mol Ther. 2016 Mar;24(3):582-91. doi: 10.1038/mt.2015.190. Epub 2015 Oct 7. Mol Ther. 2016. PMID: 26444081 Free PMC article.
-
Prototype foamy virus protease activity is essential for intraparticle reverse transcription initiation but not absolutely required for uncoating upon host cell entry.J Virol. 2013 Mar;87(6):3163-76. doi: 10.1128/JVI.02323-12. Epub 2013 Jan 2. J Virol. 2013. PMID: 23283957 Free PMC article.
References
-
- Apolonia, L., S. N. Waddington, C. Fernandes, N. J. Ward, G. Bouma, M. P. Blundell, A. J. Thrasher, M. K. Collins, and N. J. Philpott. 2007. Stable gene transfer to muscle using non-integrating lentiviral vectors. Mol. Ther. 15:1947-1954. - PubMed
-
- Butler, S. L., M. S. Hansen, and F. D. Bushman. 2001. A quantitative assay for HIV DNA integration in vivo. Nat. Med. 7:631-634. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
