

Lateralization of the central circadian pacemaker output: a test of neural control of peripheral oscillator phase
- PMID: 20592176
- PMCID: PMC2944421
- DOI: 10.1152/ajpregu.00746.2009
Lateralization of the central circadian pacemaker output: a test of neural control of peripheral oscillator phase
Abstract
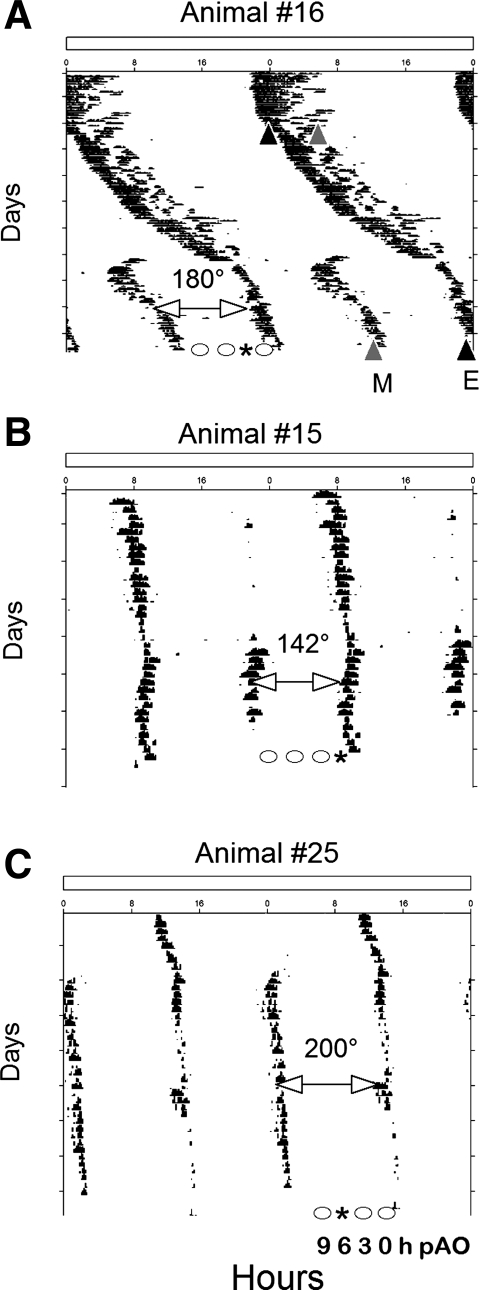
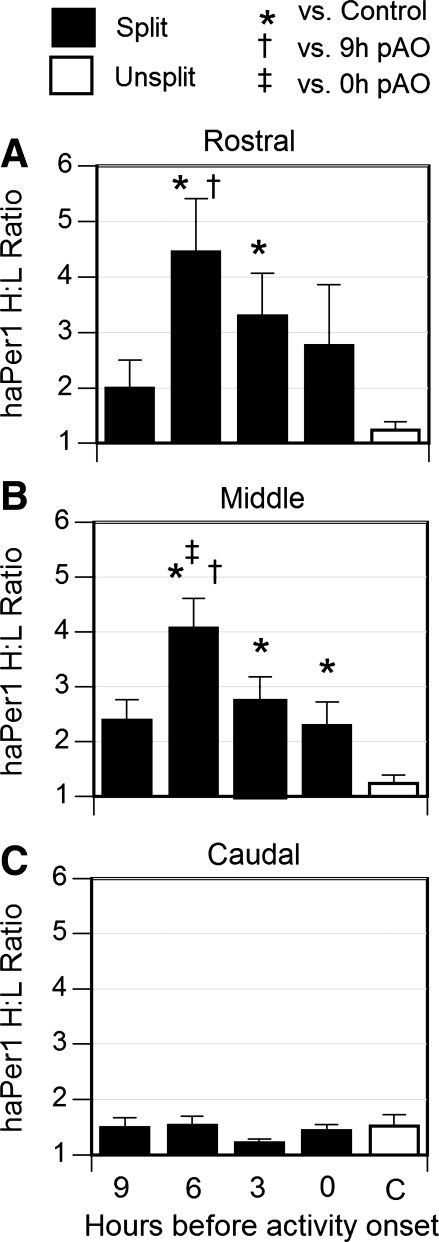
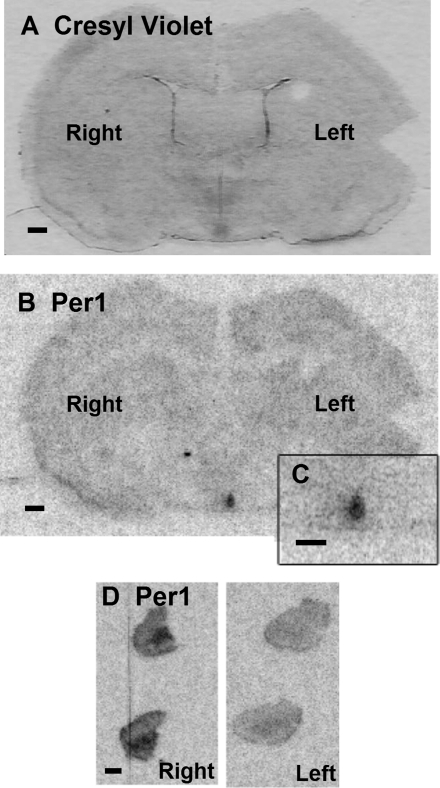
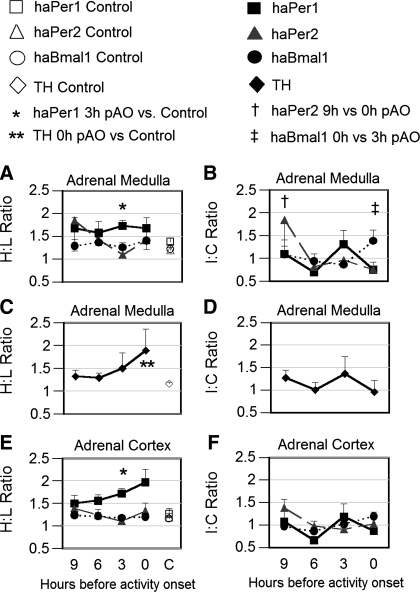
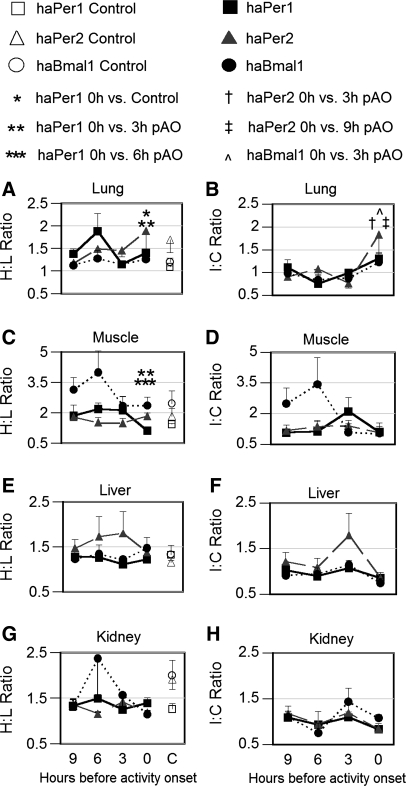
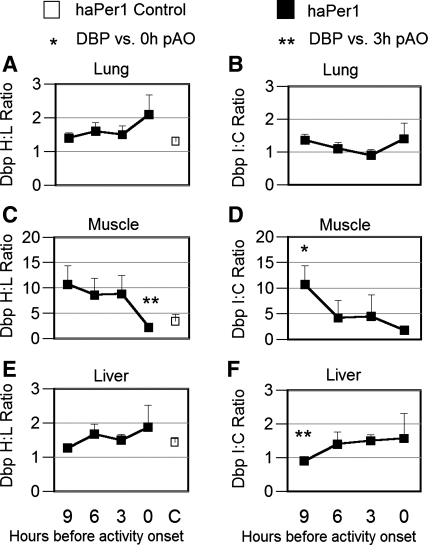
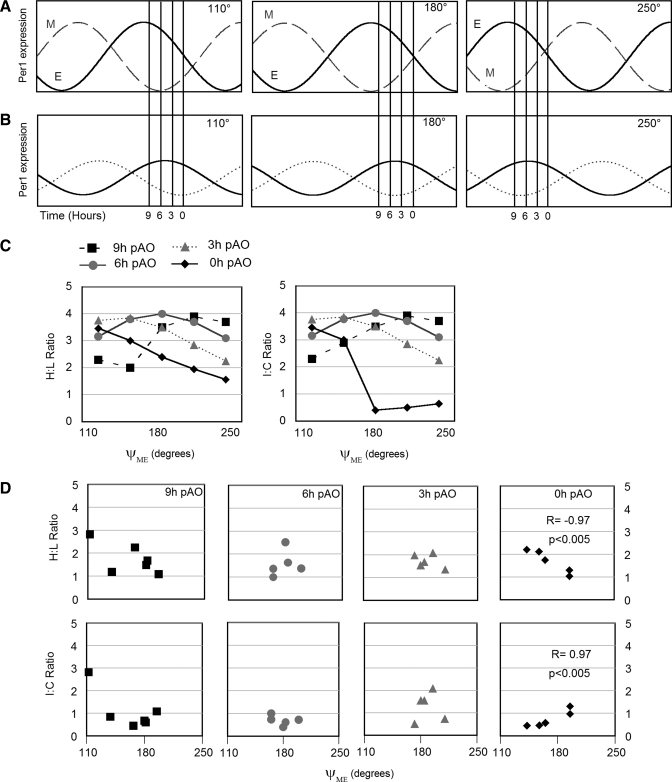
To evaluate the contribution of neural pathways to the determination of the circadian oscillator phase in peripheral organs, we assessed lateralization of clock gene expression in Syrian hamsters induced to split rhythms of locomotor activity by exposure to constant light. We measured the ratio of haPer1, haPer2, and haBmal1 mRNA on the high vs. low (H/L) side at 3-h intervals prior to the predicted activity onset (pAO). We also calculated expression on the sides ipsilateral vs. contralateral (I/C) to the side of the suprachiasmatic nucleus (SCN) expressing higher haPer1. The extent of asymmetry in split hamsters varied between specific genes, phases, and organs. Although the magnitude of asymmetry in peripheral organs was never as great as that in the SCN, we observed significantly greater lateralization of clock gene expression in the adrenal medulla and cortex, lung, and skeletal muscle, but not in liver or kidney, of split hamsters than of unsplit controls. We observed fivefold lateralization of expression of the clock-controlled gene, albumin site D-element binding protein (Dbp), in skeletal muscle (H/L: 10.7 +/- 3.7 at 3 h vs. 2.2 +/- 0.3 at 0 h pAO; P = 0.03). Furthermore, tyrosine hydroxylase expression was asymmetrical in the adrenal medulla of split (H/L: 1.9 +/- 0.5 at 0 h) vs. unsplit hamsters (1.2 +/- 0.04; P < 0.05). Consistent with a model of neurally controlled gene expression, we found significant correlations between the phase angle between morning and evening components (psi(me)) and the level of asymmetry (H/L or I/C). Our results indicate that neural pathways contribute to, but cannot completely account for, SCN regulation of the phase of peripheral oscillators.
Figures
References
-
- Abrahamson EE, Moore RY. Suprachiasmatic nucleus in the mouse: retinal innervation, intrinsic organization and efferent projections. Brain Res 916: 172–191, 2001 - PubMed
-
- Akhtar RA, Reddy AB, Maywood ES, Clayton JD, King VM, Smith AG, Gant TW, Hastings MH, Kyriacou CP. Circadian cycling of the mouse liver transcriptome, as revealed by cDNA microarray, is driven by the suprachiasmatic nucleus. Curr Biol 12: 540–550, 2002 - PubMed
-
- Balsalobre A, Brown SA, Marcacci L, Tronche F, Kellendonk C, Reichardt HM, Schütz G, Schibler U. Resetting of circadian time in peripheral tissues by glucocorticoid signaling. Science 289: 2344–2347, 2000 - PubMed
-
- Bartness TJ, Song CJ, Demas GE. SCN efferents to peripheral tissues: implications for biological rhythms. J Biol Rhythms 16: 196–204, 2001 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources