

Modeling bidirectional transcription using silkmoth chorion gene promoters
- PMID: 20592866
- PMCID: PMC2861744
- DOI: 10.4161/org.6.1.10696
Modeling bidirectional transcription using silkmoth chorion gene promoters
Abstract
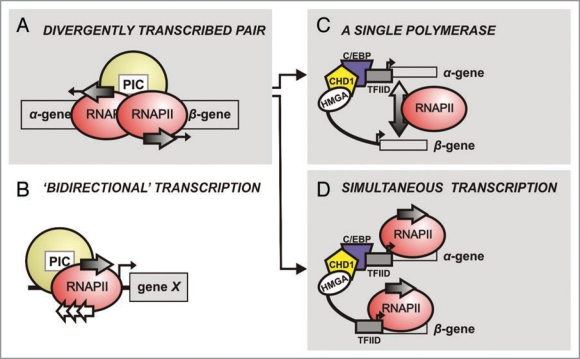
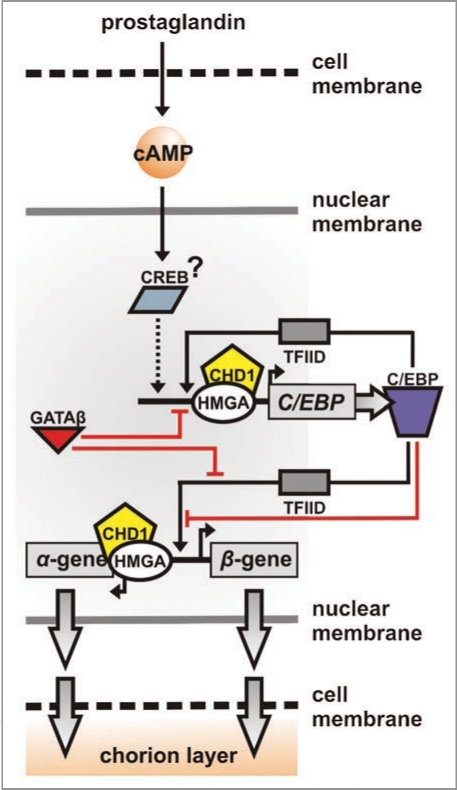
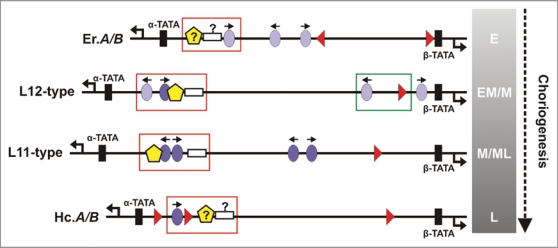
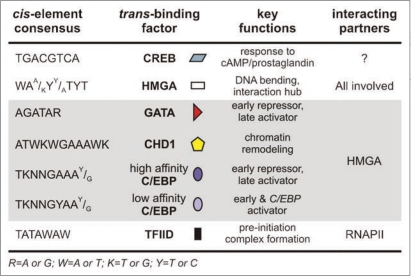
Bidirectional transcription is an interesting feature of eukaryotic genomes; yet not all aspects of its mechanism are understood. Silkmoth choriogenesis is a model system for studying transcriptional regulation at the initiation level. As chorion genes comprise a large group of divergently transcribed gene pairs, we are presented with the possibility of investigating the intricacies of bidirectional transcription. Their well characterized 5' regulatory regions and expression profiles lay the foundation for investigating protein:protein and protein:DNA interactions, and RNA polymerase function during oocyte development. In this article we summarize current knowledge on chorion gene regulation and propose an approach to modeling bidirectional transcription using chorion promoters.
Keywords: Bombyx mori; RNA polymerase II; bidirectional promoters; chorion genes; transcriptional regulation.
Figures
Similar articles
-
Analysis of developmentally regulated chorion gene promoter architecture via electroporation of silk moth follicles.Insect Mol Biol. 2015 Feb;24(1):71-81. doi: 10.1111/imb.12136. Epub 2014 Sep 25. Insect Mol Biol. 2015. PMID: 25256090
-
Gene evolution and regulation in the chorion complex of Bombyx mori. Hybridization and sequence analysis of multiple developmentally middle A/B chorion gene pairs.J Mol Biol. 1989 Sep 5;209(1):1-19. doi: 10.1016/0022-2836(89)90166-6. J Mol Biol. 1989. PMID: 2810362
-
Transgenic regulation of moth chorion gene promoters in Drosophila: tissue, temporal, and quantitative control of four bidirectional promoters.J Mol Evol. 1989 Dec;29(6):486-95. doi: 10.1007/BF02602920. J Mol Evol. 1989. PMID: 2559211
-
The ecdysone regulatory cascade and ovarian development in lepidopteran insects: insights from the silkmoth paradigm.Insect Biochem Mol Biol. 2003 Dec;33(12):1285-97. doi: 10.1016/j.ibmb.2003.06.012. Insect Biochem Mol Biol. 2003. PMID: 14599500 Review.
-
Silkmoth chorion gene regulation revisited: promoter architecture as a key player.Insect Mol Biol. 2010 Apr;19(2):141-51. doi: 10.1111/j.1365-2583.2009.00969.x. Epub 2009 Dec 7. Insect Mol Biol. 2010. PMID: 20002795 Review.
Cited by
-
Multifaceted biological insights from a draft genome sequence of the tobacco hornworm moth, Manduca sexta.Insect Biochem Mol Biol. 2016 Sep;76:118-147. doi: 10.1016/j.ibmb.2016.07.005. Epub 2016 Aug 12. Insect Biochem Mol Biol. 2016. PMID: 27522922 Free PMC article.
References
-
- Bayele HK. Trypanosoma brucei: A putative RNA polymerase II promoter. Exp Parasitol. 2009 doi:10.1016/j.exppara.2009.08.007. - PubMed
-
- Zanotto E, Häkkinen A, Teku G, Shen B, Ribeiro AS, Jacobs HT. NF-Y influences directionality of transcription from the bidirectional Mrps12/Sarsm promoter in both mouse and human cells. Biochim Biophys Acta 2. 2009;178:432–442. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials