

Macrophages as novel target cells for erythropoietin
- PMID: 20595096
- PMCID: PMC2966903
- DOI: 10.3324/haematol.2010.025015
Macrophages as novel target cells for erythropoietin
Abstract
Background: Our original demonstration of immunomodulatory effects of erythropoietin in multiple myeloma led us to the search for the cells in the immune system that are direct targets for erythropoietin. The finding that lymphocytes do not express erythropoietin receptors led to the hypothesis that other cells act as direct targets and thus mediate the effects of erythropoietin. The finding that erythropoietin has effects on dendritic cells thus led to the question of whether macrophages act as target cells for erythropoietin.
Design and methods: The effects of erythropoietin on macrophages were investigated both in-vivo and in-vitro. The in-vivo studies were performed on splenic macrophages and inflammatory peritoneal macrophages, comparing recombinant human erythropoietin-treated and untreated mice, as well as transgenic mice over-expressing human erythropoietin (tg6) and their control wild-type counterparts. The in-vitro effects of erythropoietin on macrophage surface markers and function were investigated in murine bone marrow-derived macrophages treated with recombinant human erythropoietin.
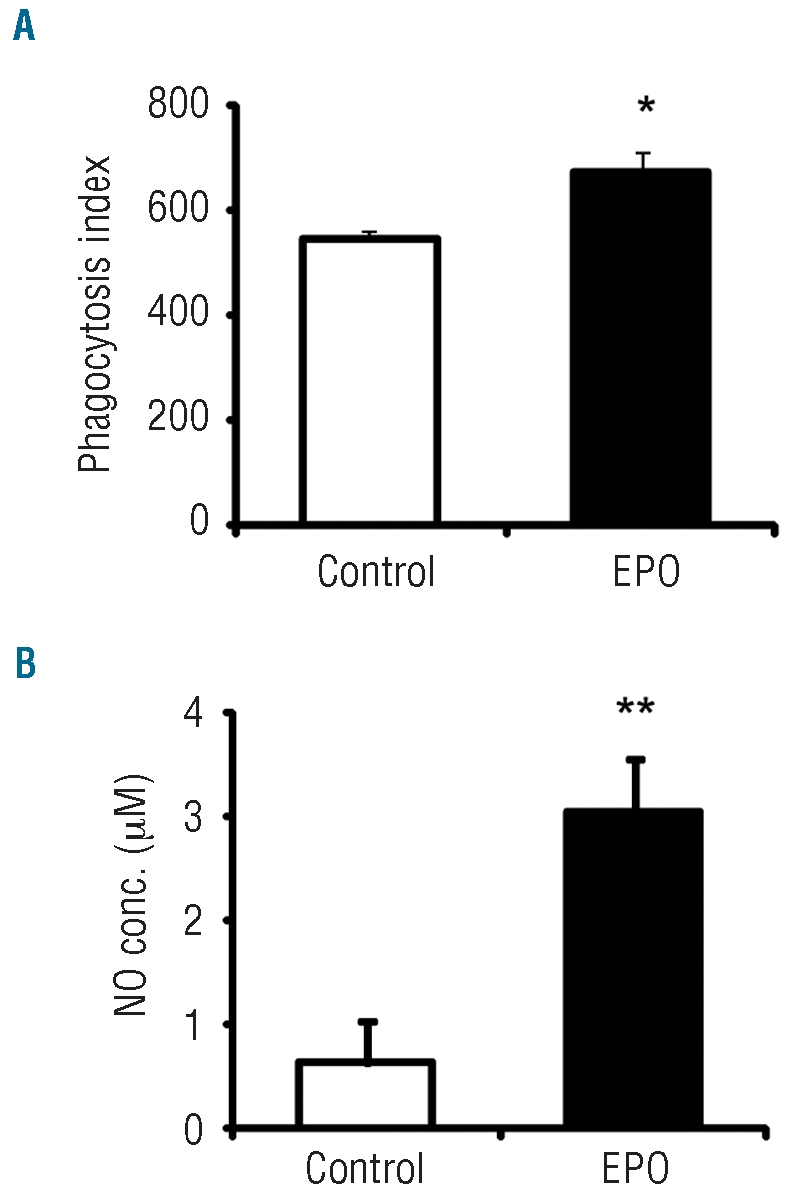
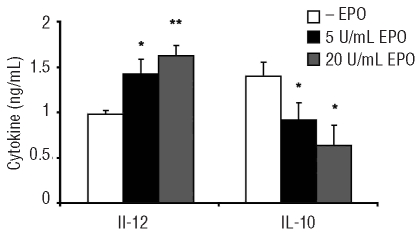
Results: Erythropoietin was found to have effects on macrophages in both the in-vivo and in-vitro experiments. In-vivo treatment led to increased numbers of splenic macrophages, and of the splenic macrophages expressing CD11b, CD80 and major histocompatibility complex class II. The peritoneal inflammatory macrophages obtained from erythropoietin-treated mice displayed increased expression of F4/80, CD11b, CD80 and major histocompatibility complex class II, and augmented phagocytic activity. The macrophages derived in-vitro from bone marrow cells expressed erythropoietin receptor transcripts, and in-vitro stimulation with erythropoietin activated multiple signaling pathways, including signal transducer and activator of transcription (STAT)1 and 5, mitogen-activated protein kinase, phosphatidylinositol 3-kinase and nuclear factor kappa B. In-vitro erythropoietin treatment of these cells up-regulated their surface expression of CD11b, F4/80 and CD80, enhanced their phagocytic activity and nitric oxide secretion, and also led to augmented interleukin 12 secretion and decreased interleukin 10 secretion in response to lipopolysaccharide.
Conclusions: Our results show that macrophages are direct targets of erythropoietin and that erythropoietin treatment enhances the pro-inflammatory activity and function of these cells. These findings point to a multifunctional role of erythropoietin and its potential clinical applications as an immunomodulating agent.
Figures




Comment in
-
Non-erythroid effects of erythropoietin.Haematologica. 2010 Nov;95(11):1803-5. doi: 10.3324/haematol.2010.030213. Haematologica. 2010. PMID: 21037325 Free PMC article. No abstract available.
-
Erythropoietin receptor is detectable on peripheral blood lymphocytes and its expression increases in activated T lymphocytes.Haematologica. 2011 Mar;96(3):e12-3; author reply e14. doi: 10.3324/haematol.2010.038414. Haematologica. 2011. PMID: 21357707 Free PMC article. No abstract available.
References
-
- Arcasoy MO. The non-haematopoietic biological effects of erythropoietin. Br J Haematol. 2008;141(1):14–31. - PubMed
-
- Mittelman M, Zeidman A, Fradin Z, Magazanik A, Lewinski UH, Cohen A. Recombinant human erythropoietin in the treatment of multiple myeloma-associated anemia. Acta Haematol. 1997;98(4):204–10. - PubMed
-
- Prutchi-Sagiv S, Golishevsky N, Oster HS, Katz O, Cohen A, Naparstek E, et al. Erythropoietin treatment in advanced multiple myeloma is associated with improved immunological functions: could it be beneficial in early disease? Br J Haematol. 2006;135(5):660–72. - PubMed
-
- Katz O, Barzilay E, Skaat A, Herman A, Mittelman M, Neumann D. Erythropoietin induced tumour mass reduction in murine lymphoproliferative models. Acta Haematol. 2005;114(3):177–9. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
