

The Prp19 complex and the Usp4Sart3 deubiquitinating enzyme control reversible ubiquitination at the spliceosome
- PMID: 20595234
- PMCID: PMC2895201
- DOI: 10.1101/gad.1925010
The Prp19 complex and the Usp4Sart3 deubiquitinating enzyme control reversible ubiquitination at the spliceosome
Abstract
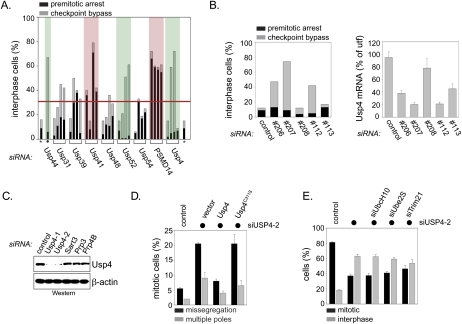
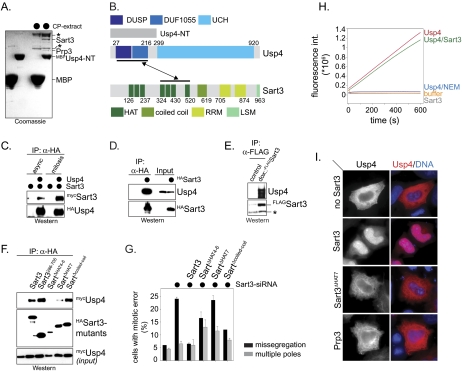
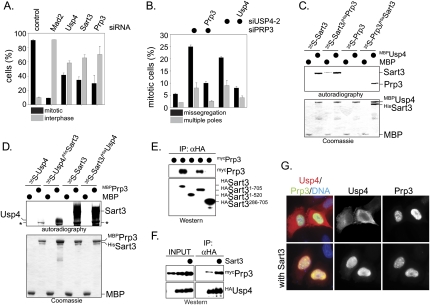
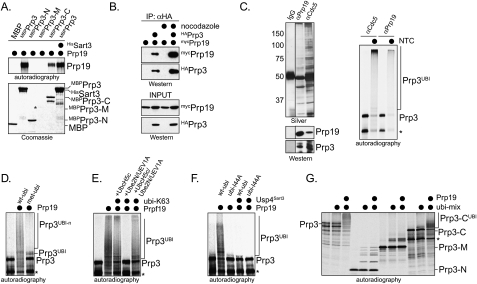
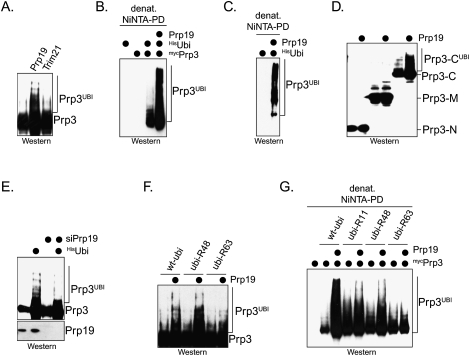
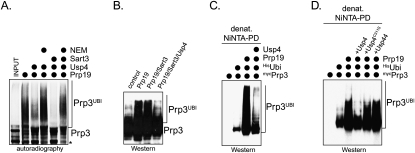
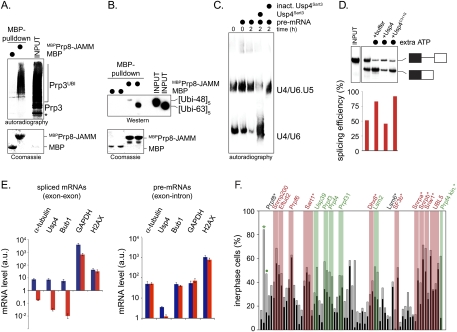
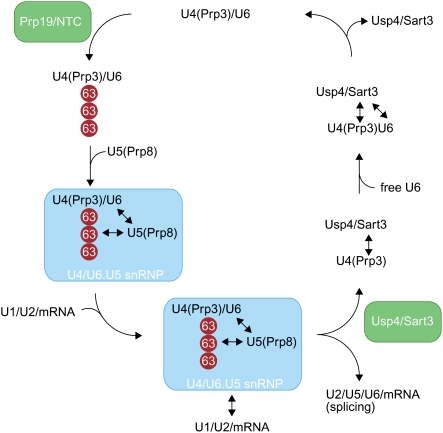
The spliceosome, a dynamic assembly of proteins and RNAs, catalyzes the excision of intron sequences from nascent mRNAs. Recent work has suggested that the activity and composition of the spliceosome are regulated by ubiquitination, but the underlying mechanisms have not been elucidated. Here, we report that the spliceosomal Prp19 complex modifies Prp3, a component of the U4 snRNP, with nonproteolytic K63-linked ubiquitin chains. The K63-linked chains increase the affinity of Prp3 for the U5 snRNP component Prp8, thereby allowing for the stabilization of the U4/U6.U5 snRNP. Prp3 is deubiquitinated by Usp4 and its substrate targeting factor, the U4/U6 recycling protein Sart3, which likely facilitates ejection of U4 proteins from the spliceosome during maturation of its active site. Loss of Usp4 in cells interferes with the accumulation of correctly spliced mRNAs, including those for alpha-tubulin and Bub1, and impairs cell cycle progression. We propose that the reversible ubiquitination of spliceosomal proteins, such as Prp3, guides rearrangements in the composition of the spliceosome at distinct steps of the splicing reaction.
Figures
Similar articles
-
When SUMO met splicing.RNA Biol. 2018;15(6):689-695. doi: 10.1080/15476286.2018.1457936. Epub 2018 May 9. RNA Biol. 2018. PMID: 29741121 Free PMC article. Review.
-
USP15 regulates dynamic protein-protein interactions of the spliceosome through deubiquitination of PRP31.Nucleic Acids Res. 2017 May 5;45(8):4866-4880. doi: 10.1093/nar/gkw1365. Nucleic Acids Res. 2017. PMID: 28088760 Free PMC article.
-
Identification of a 35S U4/U6.U5 tri-small nuclear ribonucleoprotein (tri-snRNP) complex intermediate in spliceosome assembly.J Biol Chem. 2017 Nov 3;292(44):18113-18128. doi: 10.1074/jbc.M117.797357. Epub 2017 Sep 6. J Biol Chem. 2017. PMID: 28878014 Free PMC article.
-
Recycling of the U12-type spliceosome requires p110, a component of the U6atac snRNP.Mol Cell Biol. 2004 Feb;24(4):1700-8. doi: 10.1128/MCB.24.4.1700-1708.2004. Mol Cell Biol. 2004. PMID: 14749385 Free PMC article.
-
Structural dynamics of the N-terminal domain and the Switch loop of Prp8 during spliceosome assembly and activation.Nucleic Acids Res. 2018 May 4;46(8):3833-3840. doi: 10.1093/nar/gky242. Nucleic Acids Res. 2018. PMID: 29635373 Free PMC article. Review.
Cited by
-
The increasing complexity of the ubiquitin code.Nat Cell Biol. 2016 May 27;18(6):579-86. doi: 10.1038/ncb3358. Nat Cell Biol. 2016. PMID: 27230526 Review.
-
Spotlight on USP4: Structure, Function, and Regulation.Front Cell Dev Biol. 2021 Feb 18;9:595159. doi: 10.3389/fcell.2021.595159. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33681193 Free PMC article. Review.
-
When SUMO met splicing.RNA Biol. 2018;15(6):689-695. doi: 10.1080/15476286.2018.1457936. Epub 2018 May 9. RNA Biol. 2018. PMID: 29741121 Free PMC article. Review.
-
RNA polymerase II stalling at pre-mRNA splice sites is enforced by ubiquitination of the catalytic subunit.Elife. 2017 Oct 13;6:e27082. doi: 10.7554/eLife.27082. Elife. 2017. PMID: 29027900 Free PMC article.
-
The Deubiquitinase USP4 Stabilizes Twist1 Protein to Promote Lung Cancer Cell Stemness.Cancers (Basel). 2020 Jun 15;12(6):1582. doi: 10.3390/cancers12061582. Cancers (Basel). 2020. PMID: 32549341 Free PMC article.
References
-
- Aravind L, Koonin EV 2000. The U box is a modified RING finger—A common domain in ubiquitination. Curr Biol 10: R132–R134 doi: 10.1016/S0960-9822(00)00398-5 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous