

Versatility of trigger factor interactions with ribosome-nascent chain complexes
- PMID: 20595383
- PMCID: PMC2934658
- DOI: 10.1074/jbc.M110.134163
Versatility of trigger factor interactions with ribosome-nascent chain complexes
Abstract
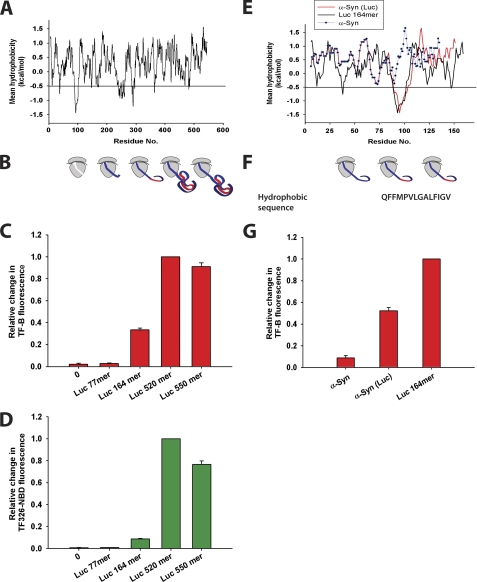
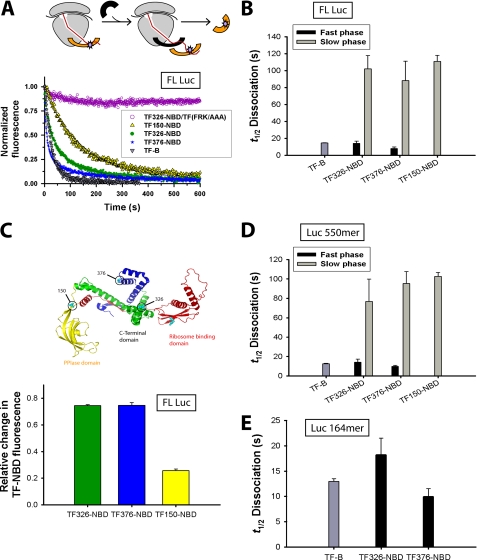
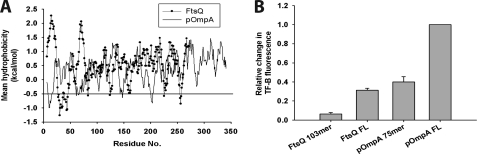
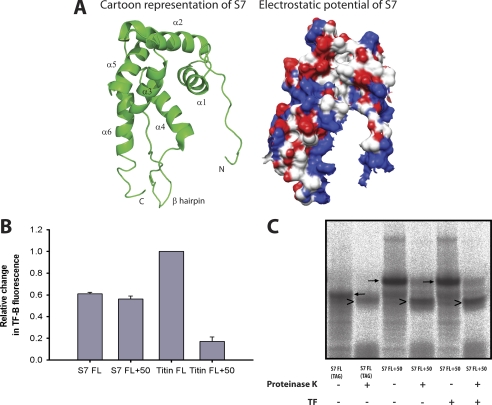
Trigger factor (TF) is the first molecular chaperone that interacts with nascent chains emerging from bacterial ribosomes. TF is a modular protein, consisting of an N-terminal ribosome binding domain, a PPIase domain, and a C-terminal domain, all of which participate in polypeptide binding. To directly monitor the interactions of TF with nascent polypeptide chains, TF variants were site-specifically labeled with an environmentally sensitive NBD fluorophore. We found a marked increase in TF-NBD fluorescence during translation of firefly luciferase (Luc) chains, which expose substantial regions of hydrophobicity, but not with nascent chains lacking extensive hydrophobic segments. TF remained associated with Luc nascent chains for 111 +/- 7 s, much longer than it remained bound to the ribosomes (t((1/2)) approximately 10-14 s). Thus, multiple TF molecules can bind per nascent chain during translation. The Escherichia coli cytosolic proteome was classified into predicted weak and strong interactors for TF, based on the occurrence of continuous hydrophobic segments in the primary sequence. The residence time of TF on the nascent chain generally correlated with the presence of hydrophobic regions and the capacity of nascent chains to bury hydrophobicity. Interestingly, TF bound the signal sequence of a secretory protein, pOmpA, but not the hydrophobic signal anchor sequence of the inner membrane protein FtsQ. On the other hand, proteins lacking linear hydrophobic segments also recruited TF, suggesting that TF can recognize hydrophobic surface features discontinuous in sequence. Moreover, TF retained significant affinity for the folded domain of the positively charged, ribosomal protein S7, indicative of an alternative mode of TF action. Thus, unlike other chaperones, TF appears to employ multiple mechanisms to interact with a wide range of substrate proteins.
Figures



Similar articles
-
Real-time observation of trigger factor function on translating ribosomes.Nature. 2006 Nov 23;444(7118):455-60. doi: 10.1038/nature05225. Epub 2006 Oct 15. Nature. 2006. PMID: 17051157
-
Interplay of signal recognition particle and trigger factor at L23 near the nascent chain exit site on the Escherichia coli ribosome.J Cell Biol. 2003 May 26;161(4):679-84. doi: 10.1083/jcb.200302130. Epub 2003 May 19. J Cell Biol. 2003. PMID: 12756233 Free PMC article.
-
Global profiling of SRP interaction with nascent polypeptides.Nature. 2016 Aug 11;536(7615):219-23. doi: 10.1038/nature19070. Epub 2016 Aug 3. Nature. 2016. PMID: 27487212
-
A cradle for new proteins: trigger factor at the ribosome.Curr Opin Struct Biol. 2005 Apr;15(2):204-12. doi: 10.1016/j.sbi.2005.03.005. Curr Opin Struct Biol. 2005. PMID: 15837180 Review.
-
Structure and function of the molecular chaperone Trigger Factor.Biochim Biophys Acta. 2010 Jun;1803(6):650-61. doi: 10.1016/j.bbamcr.2010.01.017. Epub 2010 Feb 2. Biochim Biophys Acta. 2010. PMID: 20132842 Review.
Cited by
-
Thermodynamic profiles for cotranslational trigger factor substrate recognition.Sci Adv. 2024 Jul 12;10(28):eadn4824. doi: 10.1126/sciadv.adn4824. Epub 2024 Jul 10. Sci Adv. 2024. PMID: 38985872 Free PMC article.
-
Structural characterization of the interaction of α-synuclein nascent chains with the ribosomal surface and trigger factor.Proc Natl Acad Sci U S A. 2016 May 3;113(18):5012-7. doi: 10.1073/pnas.1519124113. Epub 2016 Apr 18. Proc Natl Acad Sci U S A. 2016. PMID: 27092002 Free PMC article.
-
Mechanistic Insights into Protein Biogenesis and Maturation on the Ribosome.J Mol Biol. 2025 Feb 28:169056. doi: 10.1016/j.jmb.2025.169056. Online ahead of print. J Mol Biol. 2025. PMID: 40024436 Review.
-
Translation elongation regulates substrate selection by the signal recognition particle.J Biol Chem. 2012 Mar 2;287(10):7652-60. doi: 10.1074/jbc.M111.325001. Epub 2012 Jan 6. J Biol Chem. 2012. PMID: 22228766 Free PMC article.
-
Formyl-methionine as a degradation signal at the N-termini of bacterial proteins.Microb Cell. 2015;2(10):376-393. doi: 10.15698/mic2015.10.231. Microb Cell. 2015. PMID: 26866044 Free PMC article.
References
-
- Kramer G., Boehringer D., Ban N., Bukau B. (2009) Nat. Struct. Mol. Biol. 16, 589–597 - PubMed
-
- Hartl F. U., Hayer-Hartl M. (2009) Nat. Struct. Mol. Biol. 16, 574–581 - PubMed
-
- Hartl F. U. (1996) Nature 381, 571–579 - PubMed
-
- Frydman J., Erdjument-Bromage H., Tempst P., Hartl F. U. (1999) Nat. Struct. Biol. 6, 697–705 - PubMed
-
- Lu J., Deutsch C. (2005) Nat. Struct. Mol. Biol. 12, 1123–1129 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
