

RapGEF2 is essential for embryonic hematopoiesis but dispensable for adult hematopoiesis
- PMID: 20595512
- PMCID: PMC2974602
- DOI: 10.1182/blood-2010-01-262964
RapGEF2 is essential for embryonic hematopoiesis but dispensable for adult hematopoiesis
Abstract
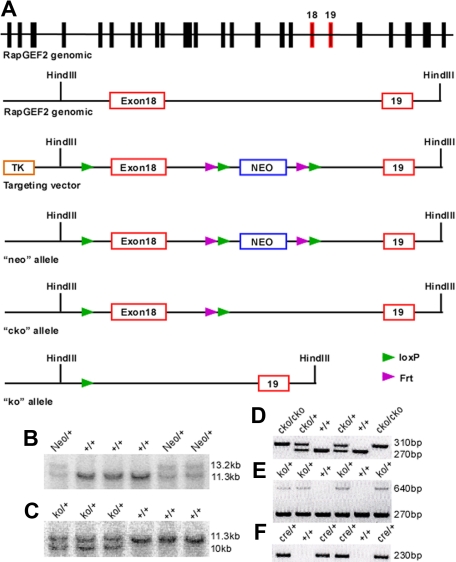
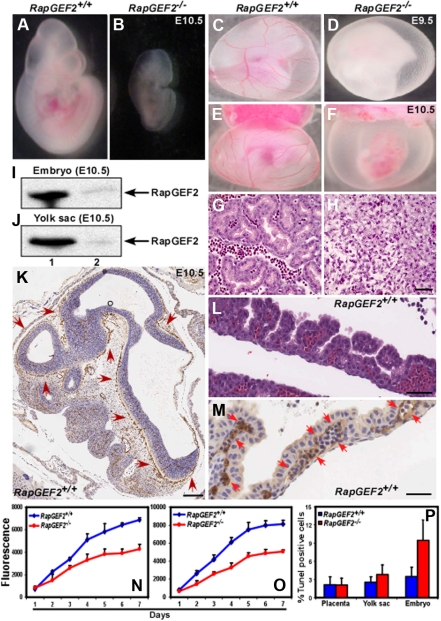
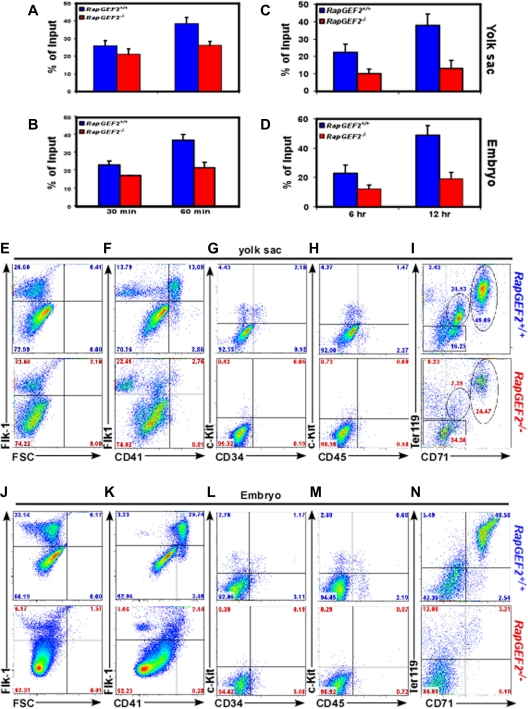
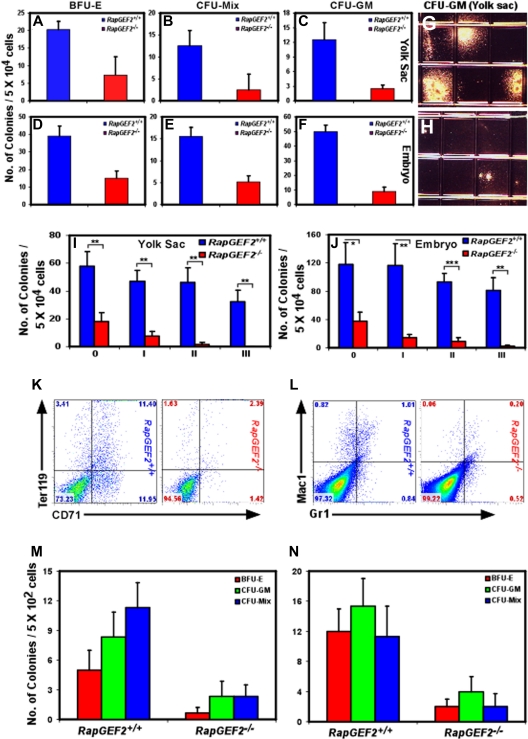
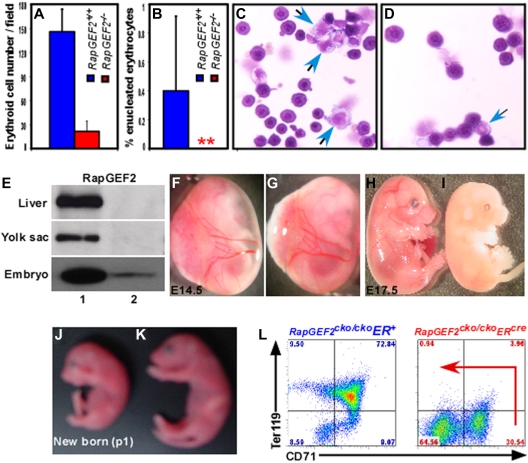
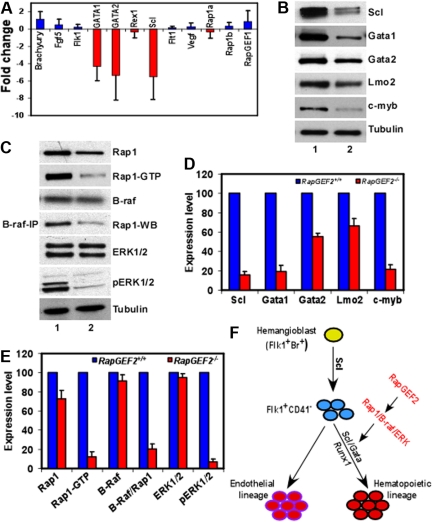
RapGEF2 is one of many guanine nucleotide exchange factors (GEFs) that specifically activate Rap1. Here, we generated RapGEF2 conditional knockout mice and studied its role in embryogenesis and fetal as well as adult hematopoietic stem cell (HSC) regulation. RapGEF2 deficiency led to embryonic lethality at ~ E11.5 due to severe yolk sac vascular defects. However, a similar number of Flk1(+) cells were present in RapGEF2(+/+) and RapGEF2(-/-) yolk sacs indicating that the bipotential early progenitors were in fact generated in the absence of RapGEF2. Further analysis of yolk sacs and embryos revealed a significant reduction of CD41 expressing cells in RapGEF2(-/-) genotype, suggesting a defect in the maintenance of definitive hematopoiesis. RapGEF2(-/-) cells displayed defects in proliferation and migration, and the in vitro colony formation ability of hematopoietic progenitors was also impaired. At the molecular level, Rap1 activation was impaired in RapGEF2(-/-) cells that in turn lead to defective B-raf/ERK signaling. Scl/Gata transcription factor expression was significantly reduced, indicating that the defects observed in RapGEF2(-/-) cells could be mediated through Scl/Gata deregulation. Inducible deletion of RapGEF2 during late embryogenesis in RapGEF2(cko/cko)ER(cre) mice leads to defective fetal liver erythropoiesis. Conversely, inducible deletion in the adult bone marrow, or specific deletion in B cells, T cells, HSCs, and endothelial cells has no impact on hematopoiesis.
Figures
Similar articles
-
Expression of CD41 marks the initiation of definitive hematopoiesis in the mouse embryo.Blood. 2003 Jan 15;101(2):508-16. doi: 10.1182/blood-2002-06-1699. Epub 2002 Sep 19. Blood. 2003. PMID: 12393529
-
The SCL +40 enhancer targets the midbrain together with primitive and definitive hematopoiesis and is regulated by SCL and GATA proteins.Mol Cell Biol. 2007 Oct;27(20):7206-19. doi: 10.1128/MCB.00931-07. Epub 2007 Aug 20. Mol Cell Biol. 2007. PMID: 17709394 Free PMC article.
-
Essential roles for Cdx in murine primitive hematopoiesis.Dev Biol. 2017 Feb 15;422(2):115-124. doi: 10.1016/j.ydbio.2017.01.002. Epub 2017 Jan 6. Dev Biol. 2017. PMID: 28065741
-
Cellular Basis of Embryonic Hematopoiesis and Its Implications in Prenatal Erythropoiesis.Int J Mol Sci. 2020 Dec 8;21(24):9346. doi: 10.3390/ijms21249346. Int J Mol Sci. 2020. PMID: 33302450 Free PMC article. Review.
-
[Hematopoietic stem cell emergence and stem cell-independent hematopoiesis in the mouse embryo].Rinsho Ketsueki. 2018;59(7):915-921. doi: 10.11406/rinketsu.59.915. Rinsho Ketsueki. 2018. PMID: 30078803 Review. Japanese.
Cited by
-
Small GTPase Rap1A/B Is Required for Lymphatic Development and Adrenomedullin-Induced Stabilization of Lymphatic Endothelial Junctions.Arterioscler Thromb Vasc Biol. 2018 Oct;38(10):2410-2422. doi: 10.1161/ATVBAHA.118.311645. Arterioscler Thromb Vasc Biol. 2018. PMID: 30354217 Free PMC article.
-
Host T Cell Dedifferentiation Effects Drive HIV-1 Latency Stability.J Virol. 2022 Mar 9;96(5):e0197421. doi: 10.1128/jvi.01974-21. Epub 2022 Jan 12. J Virol. 2022. PMID: 35019721 Free PMC article.
-
Substrate trapping proteomics reveals targets of the βTrCP2/FBXW11 ubiquitin ligase.Mol Cell Biol. 2015 Jan;35(1):167-81. doi: 10.1128/MCB.00857-14. Epub 2014 Oct 20. Mol Cell Biol. 2015. PMID: 25332235 Free PMC article.
-
Dpp dependent Hematopoietic stem cells give rise to Hh dependent blood progenitors in larval lymph gland of Drosophila.Elife. 2016 Oct 26;5:e18295. doi: 10.7554/eLife.18295. Elife. 2016. PMID: 27782877 Free PMC article.
-
Disruption of MAGI2-RapGEF2-Rap1 signaling contributes to podocyte dysfunction in congenital nephrotic syndrome caused by mutations in MAGI2.Kidney Int. 2019 Sep;96(3):642-655. doi: 10.1016/j.kint.2019.03.016. Epub 2019 Mar 28. Kidney Int. 2019. PMID: 31171376 Free PMC article.
References
-
- Pannekoek WJ, Kooistra MR, Zwartkruis FJ, Bos JL. Cell-cell junction formation: the role of Rap1 and Rap1 guanine nucleotide exchange factors. Biochim Biophys Acta. 2009;1788(4):790–796. - PubMed
-
- Retta SF, Balzac F, Avolio M. Rap1: a turnabout for the crosstalk between cadherins and integrins. Eur J Cell Biol. 2006;85(3–4):283–293. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
