

Visual sensitivity underlying changes in visual consciousness
- PMID: 20598538
- PMCID: PMC2918735
- DOI: 10.1016/j.cub.2010.06.015
Visual sensitivity underlying changes in visual consciousness
Abstract
When viewing a different stimulus with each eye, we experience the remarkable phenomenon of binocular rivalry: alternations in consciousness between the stimuli [1, 2]. According to a popular theory first proposed in 1901, neurons encoding the two stimuli engage in reciprocal inhibition [3-8] so that those processing one stimulus inhibit those processing the other, yielding consciousness of one dominant stimulus at any moment and suppressing the other. Also according to the theory, neurons encoding the dominant stimulus adapt, weakening their activity and the inhibition they can exert, whereas neurons encoding the suppressed stimulus recover from adaptation until the balance of activity reverses, triggering an alternation in consciousness. Despite its popularity, this theory has one glaring inconsistency with data: during an episode of suppression, visual sensitivity to brief probe stimuli in the dominant eye should decrease over time and should increase in the suppressed eye, yet sensitivity appears to be constant [9, 10]. Using more appropriate probe stimuli (experiment 1) in conjunction with a new method (experiment 2), we found that sensitivities in dominance and suppression do show the predicted complementary changes.
Copyright (c) 2010 Elsevier Ltd. All rights reserved.
Figures
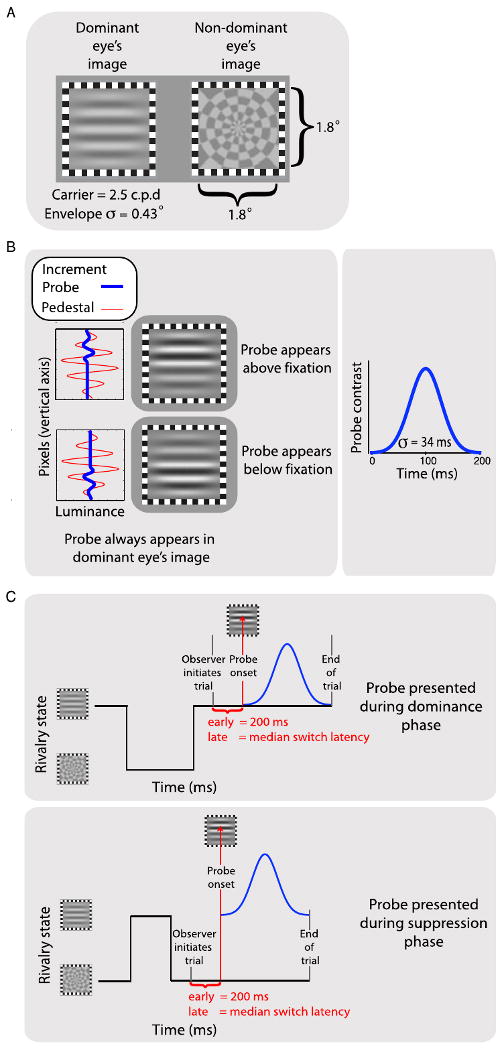
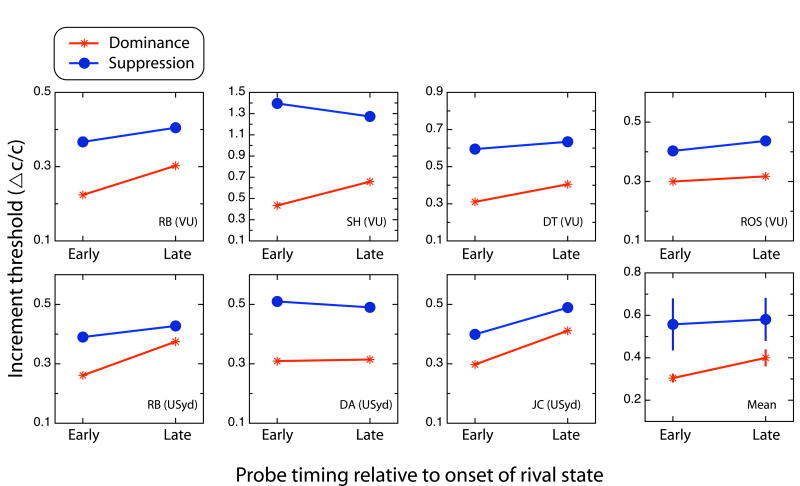
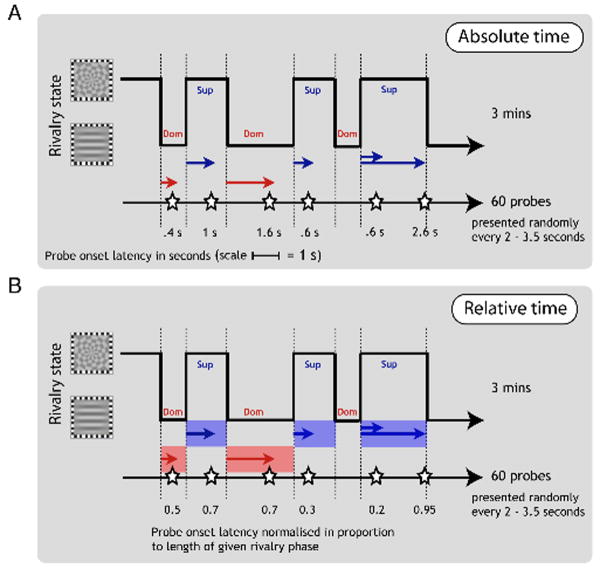
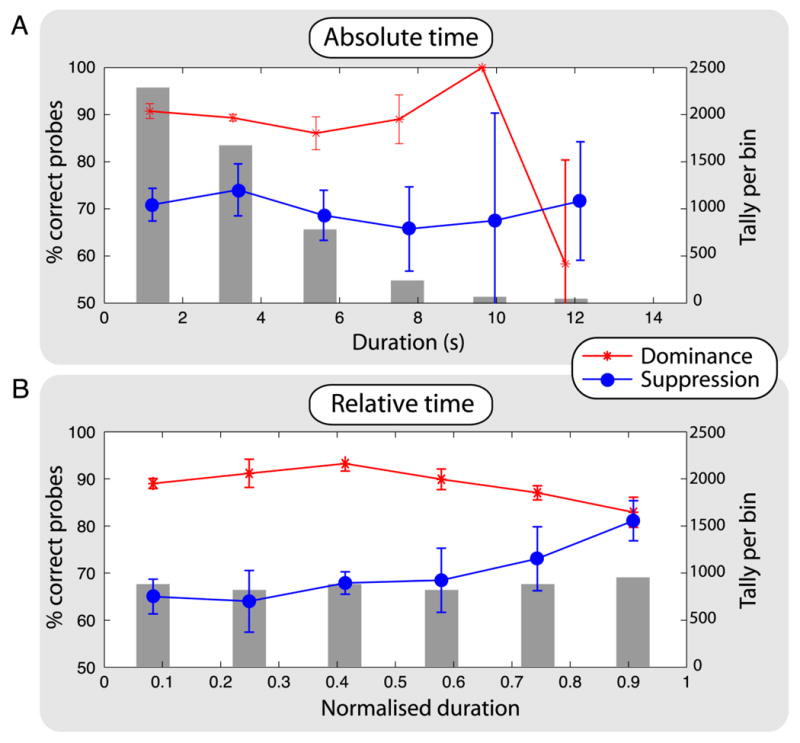
Comment in
-
Visual consciousness: the binocular rivalry explosion.Curr Biol. 2010 Aug 10;20(15):R644-6. doi: 10.1016/j.cub.2010.06.010. Curr Biol. 2010. PMID: 20692615
References
-
- Wheatstone C. Contributions to the physiology of vision.—Part the First. On some remarkable, and hitherto unobserved, phænomena of binocular vision. Philosophical Transactions of the Royal Society of London. 1838;128:371–394.
-
- Breese BB. On inhibition. Psychological Monographs. 1899;3:1–65.
-
- McDougall W. IV.—Some new observations in support of Thomas Young's theory of light- and colour-vision (I.) Mind. 1901;10:52–97.
-
- Sherrington CS. The integrative action of the nervous system. London: Constable; 1906.
-
- Sugie N. Neural models of brightness perception and retinal rivalry in binocular vision. Biological Cybernetics. 1982;43:13–21. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
