

Nonmuscle myosin II is required for cell proliferation, cell sheet adhesion and wing hair morphology during wing morphogenesis
- PMID: 20599890
- PMCID: PMC3712330
- DOI: 10.1016/j.ydbio.2010.06.028
Nonmuscle myosin II is required for cell proliferation, cell sheet adhesion and wing hair morphology during wing morphogenesis
Abstract
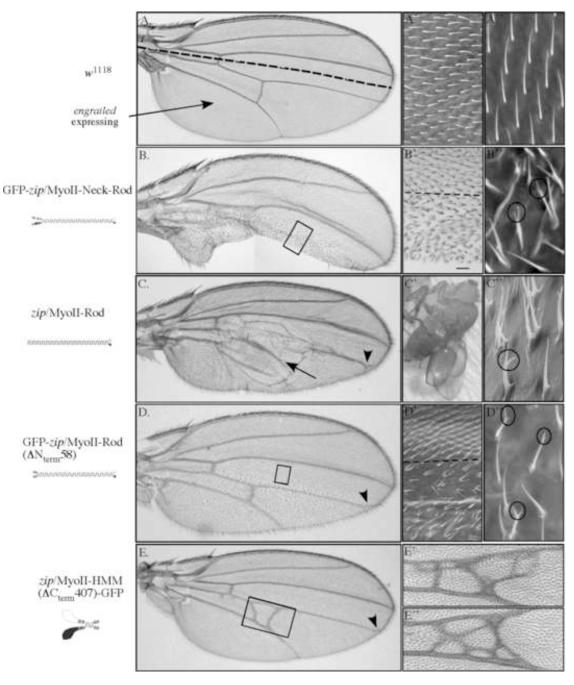
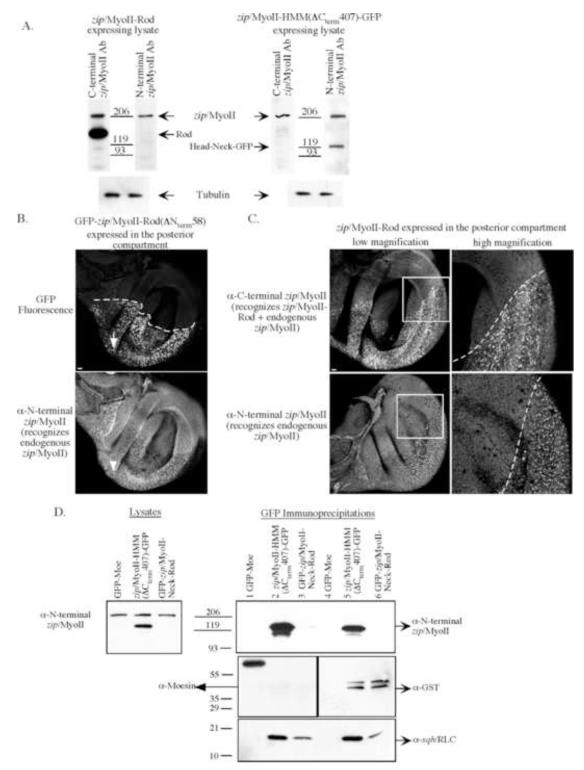
Metazoan development involves a myriad of dynamic cellular processes that require cytoskeletal function. Nonmuscle myosin II plays essential roles in embryonic development; however, knowledge of its role in post-embryonic development, even in model organisms such as Drosophila melanogaster, is only recently being revealed. In this study, truncation alleles were generated and enable the conditional perturbation, in a graded fashion, of nonmuscle myosin II function. During wing development they demonstrate novel roles for nonmuscle myosin II, including in adhesion between the dorsal and ventral wing epithelial sheets; in the formation of a single actin-based wing hair from the distal vertex of each cell; in forming unbranched wing hairs; and in the correct positioning of veins and crossveins. Many of these phenotypes overlap with those observed when clonal mosaic analysis was performed in the wing using loss of function alleles. Additional requirements for nonmuscle myosin II are in the correct formation of other actin-based cellular protrusions (microchaetae and macrochaetae). We confirm and extend genetic interaction studies to show that nonmuscle myosin II and an unconventional myosin, encoded by crinkled (ck/MyoVIIA), act antagonistically in multiple processes necessary for wing development. Lastly, we demonstrate that truncation alleles can perturb nonmuscle myosin II function via two distinct mechanisms--by titrating light chains away from endogenous heavy chains or by recruiting endogenous heavy chains into intracellular aggregates. By allowing myosin II function to be perturbed in a controlled manner, these novel tools enable the elucidation of post-embryonic roles for nonmuscle myosin II during targeted stages of fly development.
(c) 2010 Elsevier Inc. All rights reserved.
Figures





Similar articles
-
The nonmuscle myosin phosphatase PP1beta (flapwing) negatively regulates Jun N-terminal kinase in wing imaginal discs of Drosophila.Genetics. 2007 Apr;175(4):1741-9. doi: 10.1534/genetics.106.067488. Epub 2007 Feb 4. Genetics. 2007. PMID: 17277363 Free PMC article.
-
Native nonmuscle myosin II stability and light chain binding in Drosophila melanogaster.Cell Motil Cytoskeleton. 2006 Oct;63(10):604-22. doi: 10.1002/cm.20148. Cell Motil Cytoskeleton. 2006. PMID: 16917818
-
Mutations in Drosophila crinkled/Myosin VIIA disrupt denticle morphogenesis.Dev Biol. 2021 Feb;470:121-135. doi: 10.1016/j.ydbio.2020.11.007. Epub 2020 Nov 25. Dev Biol. 2021. PMID: 33248112 Free PMC article.
-
Mammalian nonmuscle myosin II comes in three flavors.Biochem Biophys Res Commun. 2018 Nov 25;506(2):394-402. doi: 10.1016/j.bbrc.2018.03.103. Epub 2018 Mar 17. Biochem Biophys Res Commun. 2018. PMID: 29550471 Free PMC article. Review.
-
The PS integrins are required for a regulatory event during Drosophila wing morphogenesis.Ann N Y Acad Sci. 1998 Oct 23;857:99-109. doi: 10.1111/j.1749-6632.1998.tb10110.x. Ann N Y Acad Sci. 1998. PMID: 9917835 Review.
Cited by
-
Systematic analysis of the physiological importance of deubiquitinating enzymes.PLoS One. 2012;7(8):e43112. doi: 10.1371/journal.pone.0043112. Epub 2012 Aug 24. PLoS One. 2012. PMID: 22937016 Free PMC article.
-
Drosophila as a Model of Unconventional Translation in Spinocerebellar Ataxia Type 3.Cells. 2022 Apr 4;11(7):1223. doi: 10.3390/cells11071223. Cells. 2022. PMID: 35406787 Free PMC article.
-
Genome-Wide Identification, Characterization and Expression Profiling of myosin Family Genes in Sebastes schlegelii.Genes (Basel). 2021 May 25;12(6):808. doi: 10.3390/genes12060808. Genes (Basel). 2021. PMID: 34070681 Free PMC article.
-
Regulation of Notch signaling by non-muscle myosin II Zipper in Drosophila.Cell Mol Life Sci. 2024 Apr 24;81(1):195. doi: 10.1007/s00018-024-05142-1. Cell Mol Life Sci. 2024. PMID: 38653877 Free PMC article.
-
Kinetic characterization of the sole nonmuscle myosin-2 from the model organism Drosophila melanogaster.FASEB J. 2015 Apr;29(4):1456-66. doi: 10.1096/fj.14-266742. Epub 2015 Jan 30. FASEB J. 2015. PMID: 25636739 Free PMC article.
References
-
- Adler PN. Planar signaling and morphogenesis in Drosophila. Dev Cell. 2002;2:525–35. - PubMed
-
- Adler PN, et al. Tissue polarity points from cells that have higher Frizzled levels towards cells that have lower Frizzled levels. Curr Biol. 1997;7:940–9. - PubMed
-
- Barros CS, et al. Drosophila nonmuscle myosin II promotes the asymmetric segregation of cell fate determinants by cortical exclusion rather than active transport. Dev Cell. 2003;5:829–40. - PubMed
-
- Bertet C, et al. Myosin-dependent junction remodelling controls planar cell intercalation and axis elongation. Nature. 2004;429:667–71. - PubMed
-
- Blair SS. Wing vein patterning in Drosophila and the analysis of intercellular signaling. Annu Rev Cell Dev Biol. 2007;23:293–319. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
