

Galectin-3: A novel substrate for c-Abl kinase
- PMID: 20600357
- PMCID: PMC2923841
- DOI: 10.1016/j.bbamcr.2010.06.007
Galectin-3: A novel substrate for c-Abl kinase
Abstract
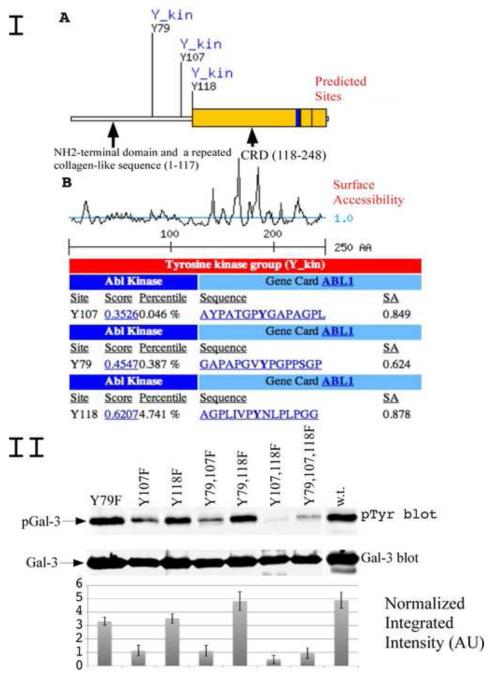
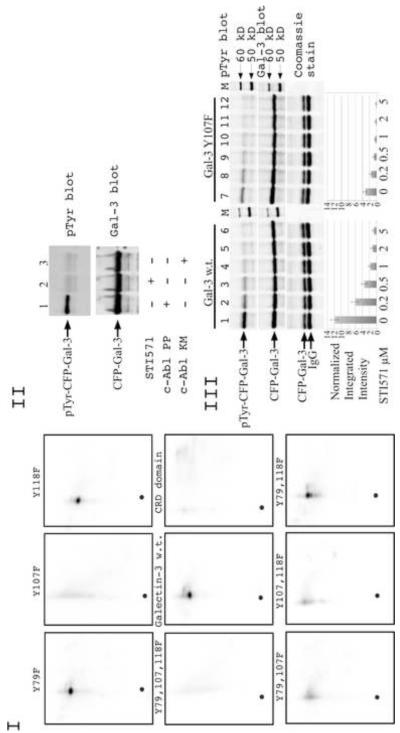
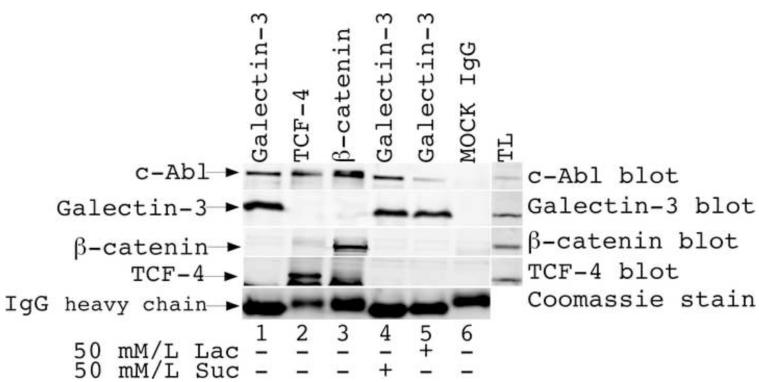
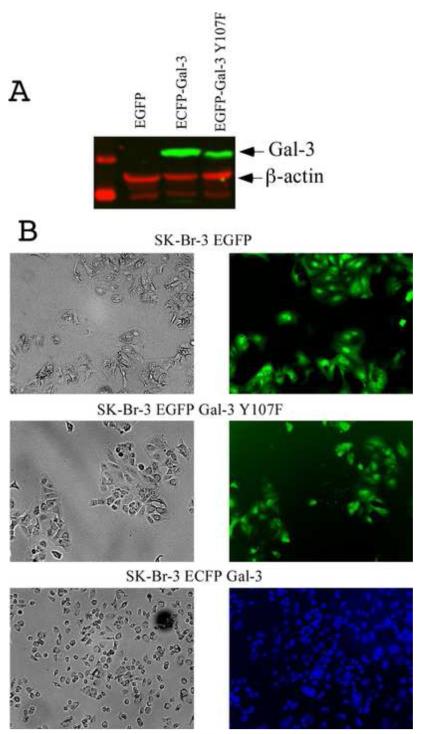
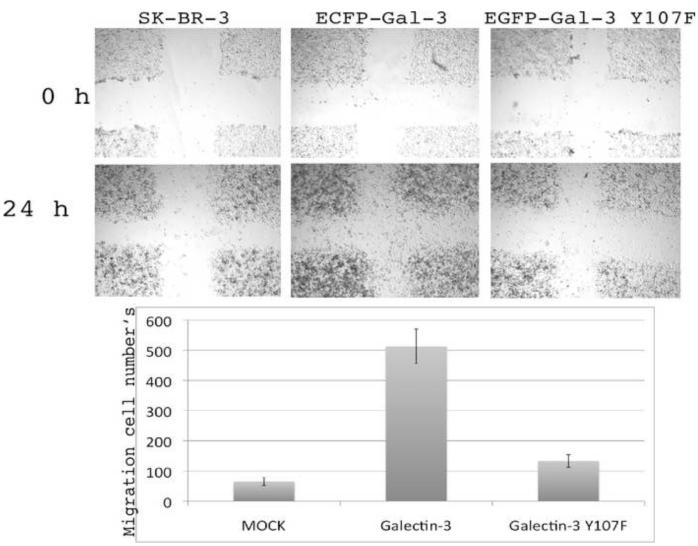
Galectin-3, a beta-galactoside-binding lectin, is found in cellular and extracellular location of the cell and has pleiotropic biological functions such as cell growth, cell adhesion and cell-cell interaction. It may exhibit anti- or pro-apoptotic activity depending on its localization and post-translational modifications. Two important post-translational modifications of galectin-3 have been reported: its cleavage and phosphorylation. Cleavage of galectin-3 was reported to be involved with angiogenic potential and apoptotic resistance. Phosphorylation of galectin-3 regulates its sugar-binding ability. In this report we have identified novel tyrosine phosphorylation sites in galectin-3 as well as the kinase responsible for its phosphorylation. Our results demonstrate that tyrosines at positions 79, 107 and 118 can be phosphorylated in vitro and in vivo by c-Abl kinase. Tyrosine 107 is the main target of c-Abl. Expression of galectin-3 Y107F mutant in galectin-3 null SK-Br-3 cells leads to morphological changes and increased motility compared to wild type galectin-3. Further investigation is needed to better understand the functional significance of the novel tyrosine phosphorylated sites of galectin-3.
Copyright © 2010 Elsevier B.V. All rights reserved.
Figures
References
-
- Barondes SH, Castronovo V, Cooper DN, Cummings RD, Drickamer K, Feizi T, Gitt MA, Hirabayashi J, Hughes C, Kasai K. Galectins: a family of animal beta-galactoside-binding lectins. Cell. 1994;76:597–598. - PubMed
-
- Barondes SH, Cooper DN, Gitt MA, Leffler H. Galectins. Structure and function of a large family of animal lectins. J. Biol. Chem. 1994;269:20807–20810. - PubMed
-
- Califice S, Castronovo V, Van Den Brule F. Galectin-3 and cancer. Int. J. Oncol. 2004;25:983–992. - PubMed
-
- Dumic J, Dabelic S, Flogel M. Galectin-3: an open-ended story. Biochim. Biophys. Acta. 2006;1760:616–635. - PubMed
-
- Huflejt ME, Turck CW, Lindstedt R, Barondes SH, Leffler H. L-29, a soluble lactose-binding lectin, is phosphorylated on serine 6 and serine 12 in vivo and by casein kinase I. J. Biol. Chem. 1993;268:26712–26718. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
