

doi: 10.1098/rstb.2009.0322.
Shining light into the black box of spinal locomotor networks
Affiliations
- PMID: 20603359
- PMCID: PMC2894950
- DOI: 10.1098/rstb.2009.0322
Item in Clipboard
Shining light into the black box of spinal locomotor networks
Philos Trans R Soc Lond B Biol Sci.
.
Abstract
Rhythmic activity is responsible for numerous essential motor functions including locomotion, breathing and chewing. In the case of locomotion, it has been realized for some time that the spinal cord contains sufficient circuitry to produce a sophisticated stepping pattern. However, the central pattern generator for locomotion in mammals has remained a 'black box' where inputs to the network were manipulated and the outputs interpreted. Over the last decade, new genetic approaches and techniques have been developed that provide ways to identify and manipulate the activity of classes of interneurons. The use of these techniques will be critically discussed and related to current models of network function.
Figures
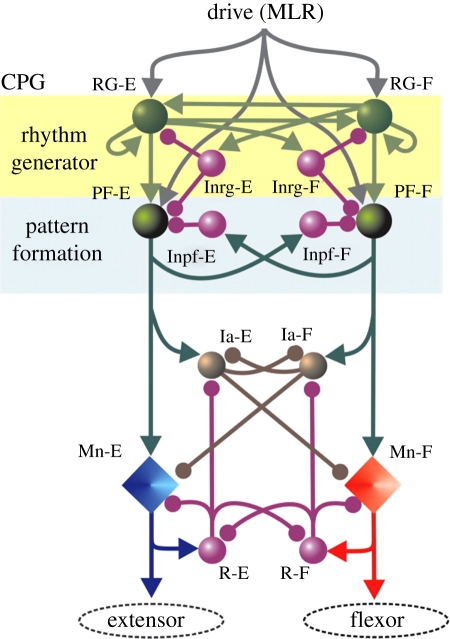
Schematic of model by Rybak & McCrea. The populations of interneurons are indicated by spheres, while the motoneurons are represented by diamonds. This three-layer model consists of a rhythm-generating layer of extensor (RG-E) and flexor (RG-F) interneurons. Both populations have recurrent excitatory connections (see also figure 2). These interneurons in turn receive mutually inhibitory input (Inrg cells). The drive projects to a pattern formation layer (PF), which acts through mutually inhibitory connections (Inpf cells) to sculpt the pattern, which is then output to the extensor and flexor motoneurons. The final output of the motoneurons is modulated by a final layer of Ia inhibitory interneurons (Ia-E, Ia-F) and Renshaw cells (R-E, R-F). Arrows indicate excitatory drive, while the filled circles indicate inhibitory drive. Reproduced with permission.
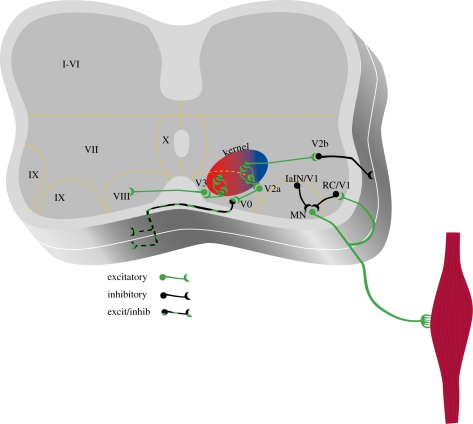
Genetic techniques can be used to dissect circuits within the spinal cord. Populations of interneurons derived from progenitor cells can be identified in neonatal mice. Schematic shows different populations of cells that can be identified currently in neonatal mouse preparations. V0 cells are CCINs that consist of both glutamatergic and glycinergic/GABAergic cells. Animals lacking these commissural cells have poor left–right coordination. V1 interneurons are considered to be inhibitory and are a heterogeneous population. At least a portion of them is involved in regulating the speed of the rhythm. Renshaw cells and Ia inhibitory interneurons (IaINs) form part of the V1 population and have been well characterized, but the contribution of individual classes of V1 neurons to rhythm generation has not been well established. V2 cells are also heterogeneous and consist of a population of excitatory interneurons (V2a) that project ipsilaterally and are involved in the regulation of left–right coordination. Another population of inhibitory interneurons possibly projects caudally and regulates flexor–extensor alternation. V3 cells are glutamatergic CCINs that may play a role in coordination of locomotor patterns. Interneurons that belong to the kernel that generates the timing pattern have not been conclusively identified. See text for references.
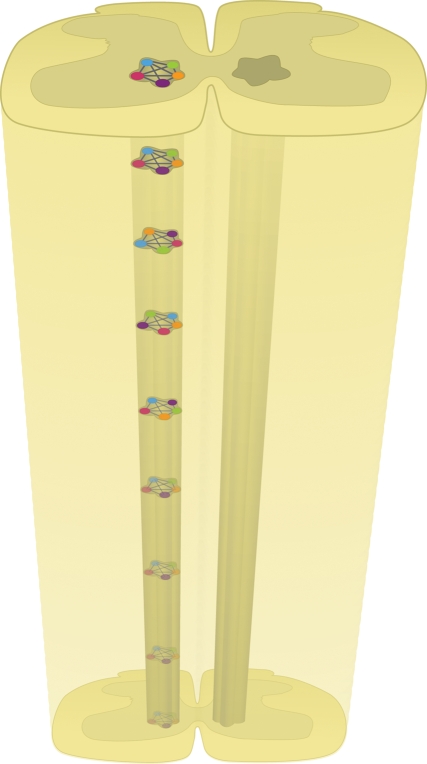
The rhythm generator kernel is probably distributed throughout the spinal cord. This diagram shows different classes of mutually excitatory populations of interneurons that form one part of a rhythm generator (RG) half-centre. The different classes comprising the single RG can be interconnected in different ways throughout the network. In this system, deletion of a single class of interneurons would not be expected to completely block the rhythm.
References
-
- Airan R. D., Hu E. S., Vijaykumar R., Roy M., Meltzer L. A., Deisseroth K.2007Integration of light-controlled neuronal firing and fast circuit imaging. Curr. Opin. Neurobiol. 17, 587–592 (doi:10.1016/j.conb.2007.11.003) - DOI - PMC - PubMed
-
- Al-Mosawie A., Wilson J. M., Brownstone R. M.2007Heterogeneity of V2-derived interneurons in the adult mouse spinal cord. Eur. J. Neurosci. 26, 3003–3015 (doi:10.1111/j.1460-9568.2007.05907.x) - DOI - PubMed
-
- Alvarez F. J., Jonas P. C., Sapir T., Hartley R., Berrocal M. C., Geiman E. J., Todd A. J., Goulding M.2005Postnatal phenotype and localization of spinal cord V1 derived interneurons. J. Comp. Neurol. 493, 177–192 (doi:10.1002/cne.20711) - DOI - PMC - PubMed
-
- Bannatyne B. A., Edgley S. A., Hammar I., Jankowska E., Maxwell D. J.2003Networks of inhibitory and excitatory commissural interneurons mediating crossed reticulospinal actions. Eur. J. Neurosci. 18, 2273–2284 (doi:10.1046/j.1460-9568.2003.02973.x) - DOI - PMC - PubMed
-
- Barbeau H., Rossignol S.1987Recovery of locomotion after chronic spinalization in the adult cat. Brain Res. 412, 84–95 (doi:10.1016/0006-8993(87)91442-9) - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources