

Ubiquitination in postsynaptic function and plasticity
- PMID: 20604708
- PMCID: PMC3163670
- DOI: 10.1146/annurev-cellbio-100109-104129
Ubiquitination in postsynaptic function and plasticity
Abstract
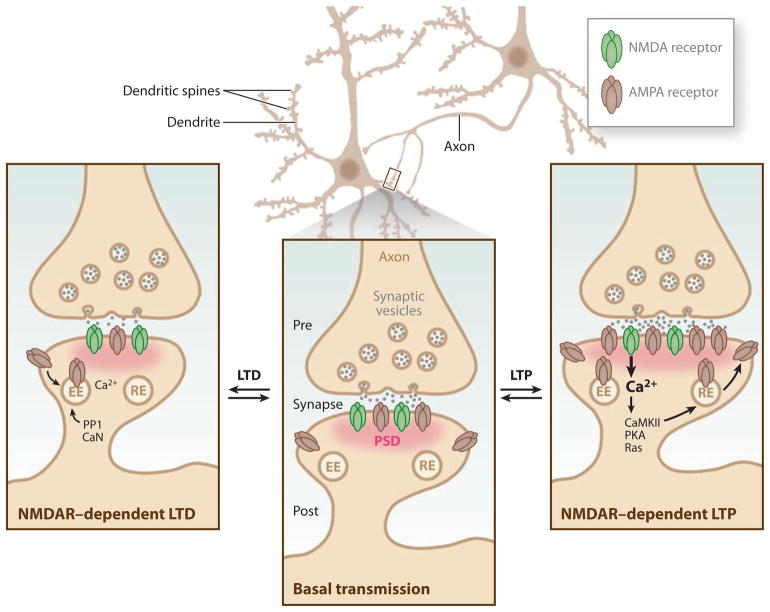
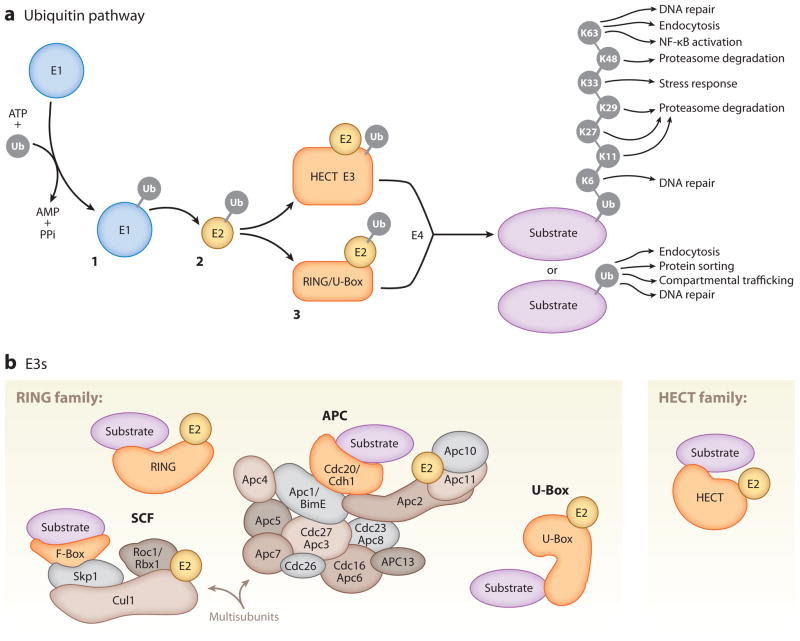
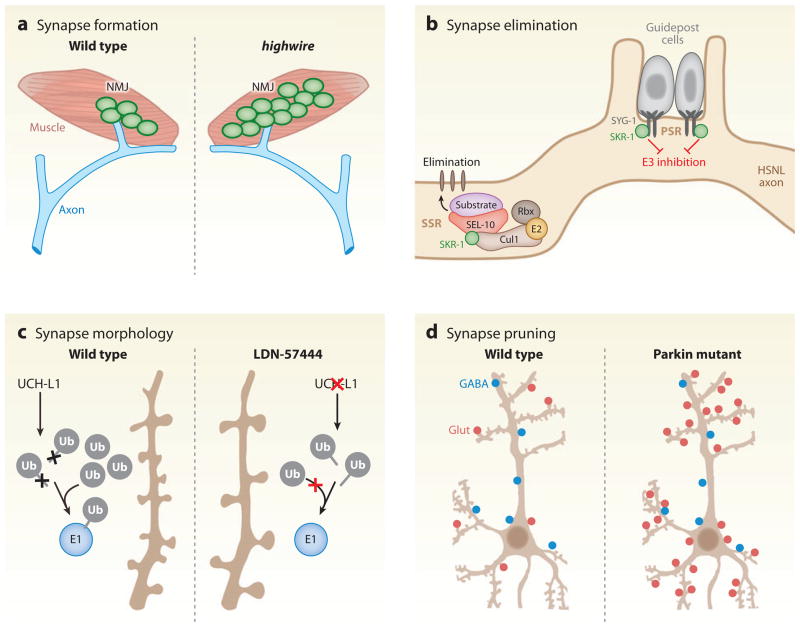
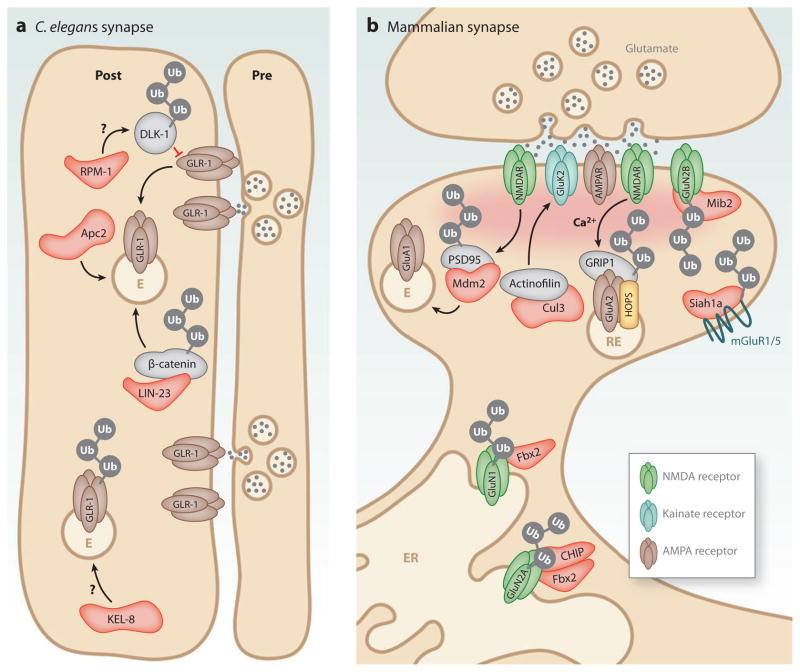
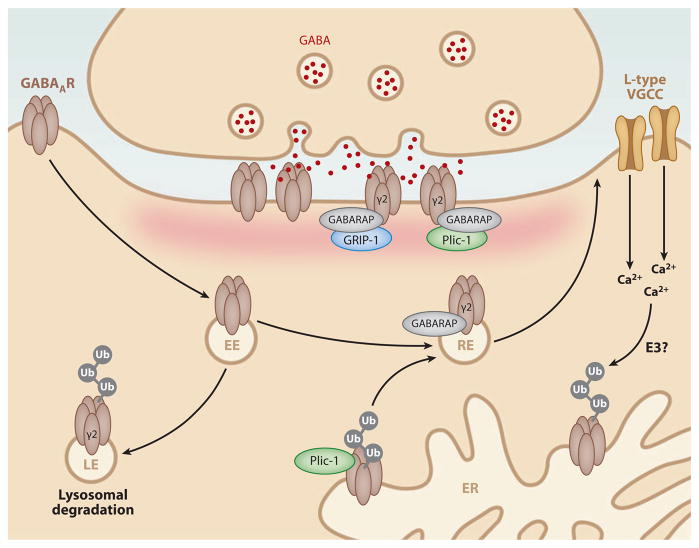
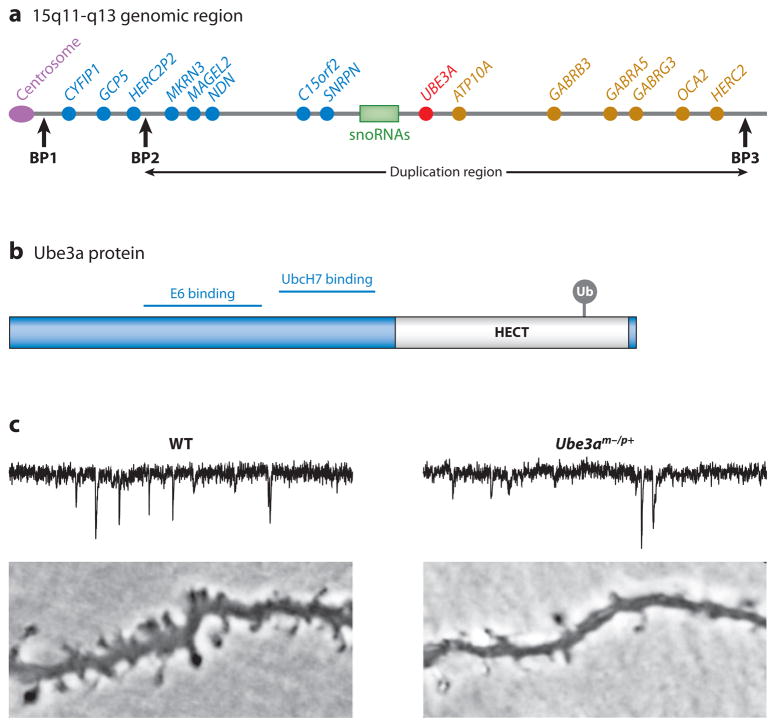
Neurons are highly specialized cells whose connectivity at synapses subserves rapid information transfer in the brain. Proper information processing, learning, and memory storage in the brain requires continuous remodeling of synaptic networks. Such remodeling includes synapse formation, elimination, synaptic protein turnover, and changes in synaptic transmission. An emergent mechanism for regulating synapse function is posttranslational modification through the ubiquitin pathway at the postsynaptic membrane. Here, we discuss recent findings implicating ubiquitination and protein degradation in postsynaptic function and plasticity. We describe postsynaptic ubiquitination pathways and their role in brain development, neuronal physiology, and brain disorders.
Figures
References
-
- Ahmari SE, Buchanan J, Smith SJ. Assembly of presynaptic active zones from cytoplasmic transport packets. Nat Neurosci. 2000;3:445–51. - PubMed
-
- Anderson C, Crimmins S, Wilson JA, Korbel GA, Ploegh HL, Wilson SM. Loss of Usp14 results in reduced levels of ubiquitin in ataxia mice. J Neurochem. 2005;95:724–31. - PubMed
-
- Artinian J, McGauran AM, De Jaeger X, Mouledous L, Frances B, Roullet P. Protein degradation, as with protein synthesis, is required during not only long-term spatial memory consolidation but also reconsolidation. Eur J Neurosci. 2008;27:3009–19. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
