

The levels of male gametic mitochondrial DNA are highly regulated in angiosperms with regard to mitochondrial inheritance
- PMID: 20605854
- PMCID: PMC2929101
- DOI: 10.1105/tpc.109.071902
The levels of male gametic mitochondrial DNA are highly regulated in angiosperms with regard to mitochondrial inheritance
Abstract
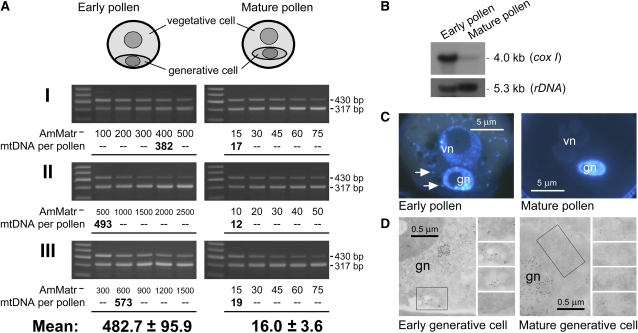
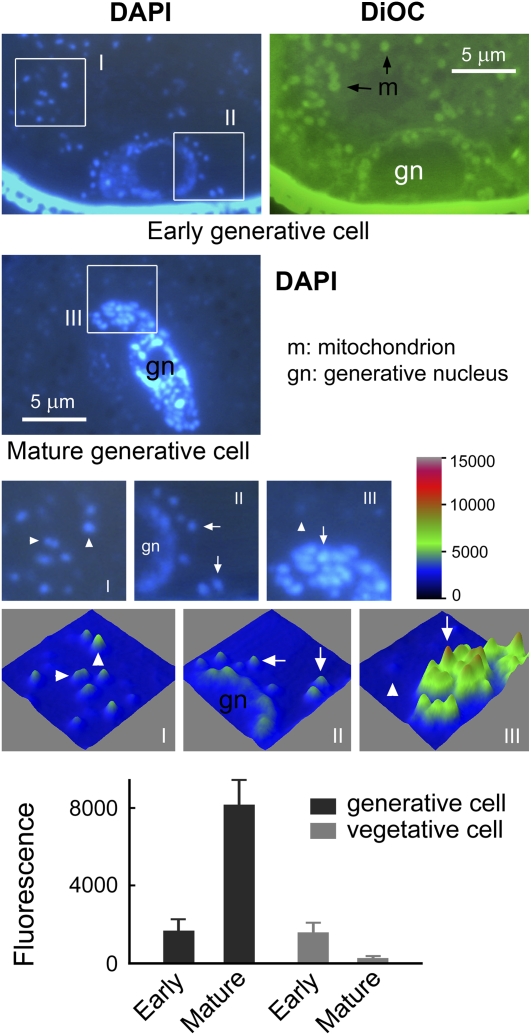
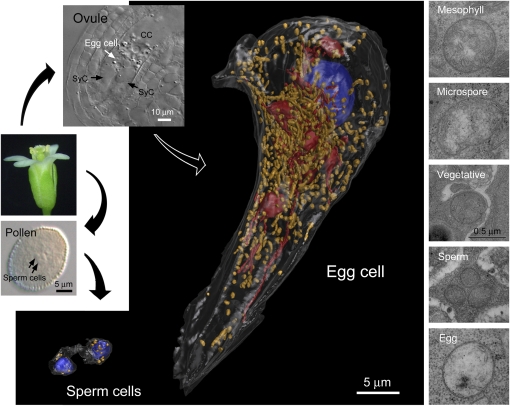
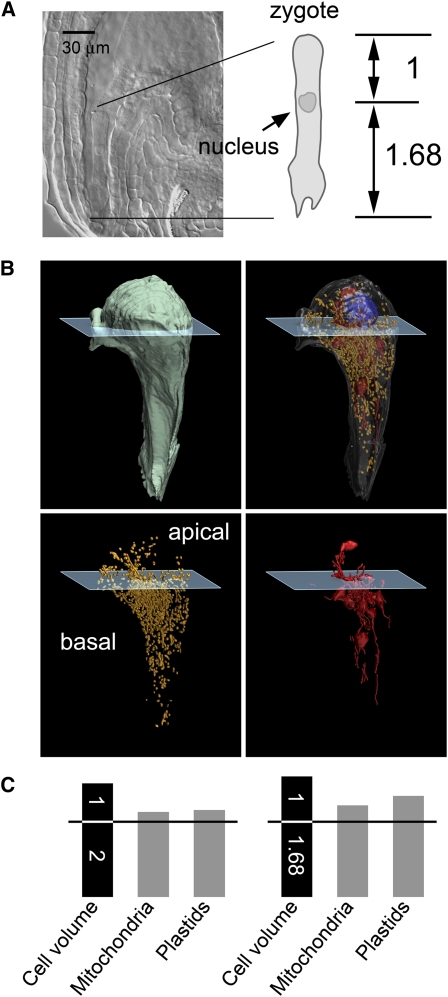
The mechanisms that regulate mitochondrial inheritance are not yet clear, even though it is 100 years since the first description of non-Mendelian genetics. Here, we quantified the copy numbers of mitochondrial DNA (mtDNA) in the gametic cells of angiosperm species. We demonstrate that each egg cell from Arabidopsis thaliana, Antirrhinum majus, and Nicotiana tabacum possesses 59.0, 42.7, and 73.0 copies of mtDNA on average, respectively. These values are equivalent to those in Arabidopsis mesophyll cells, at 61.7 copies per cell. On the other hand, sperm or generative cells from Arabidopsis, A. majus, and N. tabacum possess minor amounts of mtDNA, at 0.083, 0.47, and 1 copy on average, respectively. We further reveal a 50-fold degradation of mtDNA during pollen development in A. majus. In contrast, markedly high levels of mtDNA are found in the male gametic cells of Cucumis melo and Pelargonium zonale (1296.3 and 256.7 copies, respectively). Our results provide direct evidence for mitochondrial genomic insufficiency in the eggs and somatic cells and indicate that a male gamete of an angiosperm may possess mtDNA at concentrations as high as 21-fold (C. melo) or as low as 0.1% (Arabidopsis) of the levels in somatic cells. These observations reveal the existence of a strong regulatory system for the male gametic mtDNA levels in angiosperms with regard to mitochondrial inheritance.
Figures
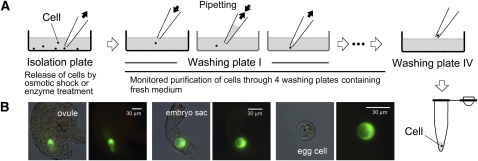
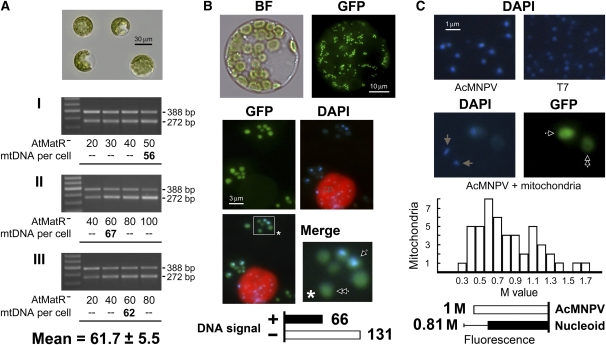
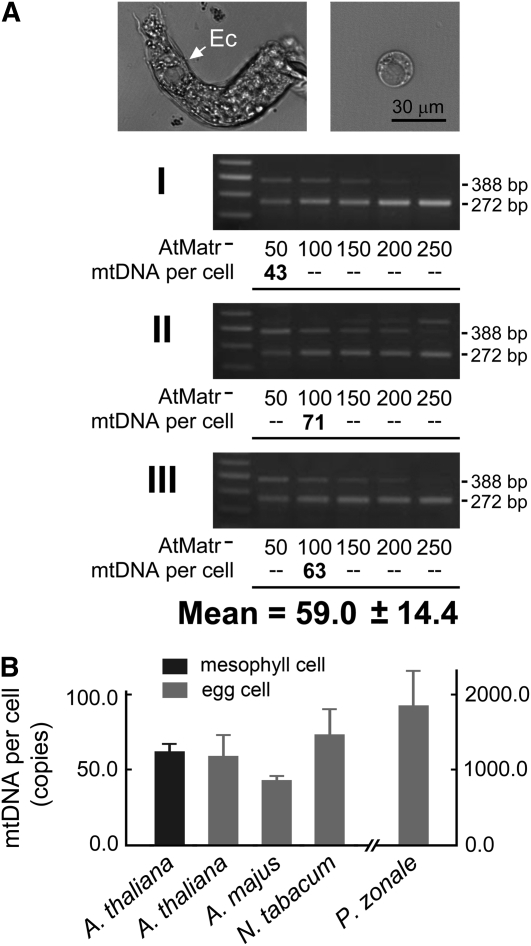
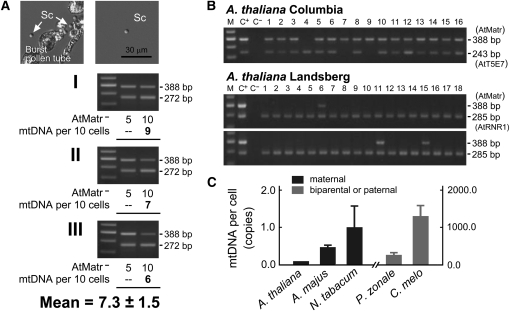
References
-
- Baur E. (1909). Das Wesen und die Erblichkeitsverhältnisse der “Varietates albomarginatae hort.” von Pelargonium zonale Z. Indukt. Abstammungs-Vererbungsl. 1: 330–351
-
- Bendich A.J., Gauriloff L.P. (1984). Morphometric analysis of Cucurbit mitochondria: The relationship between chondriome volume and DNA content. Protoplasma 119: 1–7
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
