

A flexible RNA backbone within the polypyrimidine tract is required for U2AF65 binding and pre-mRNA splicing in vivo
- PMID: 20606010
- PMCID: PMC2937558
- DOI: 10.1128/MCB.00531-10
A flexible RNA backbone within the polypyrimidine tract is required for U2AF65 binding and pre-mRNA splicing in vivo
Abstract
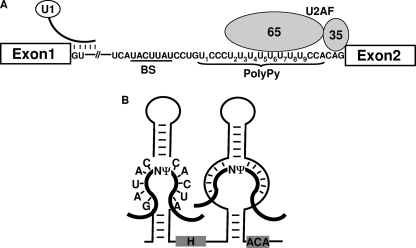
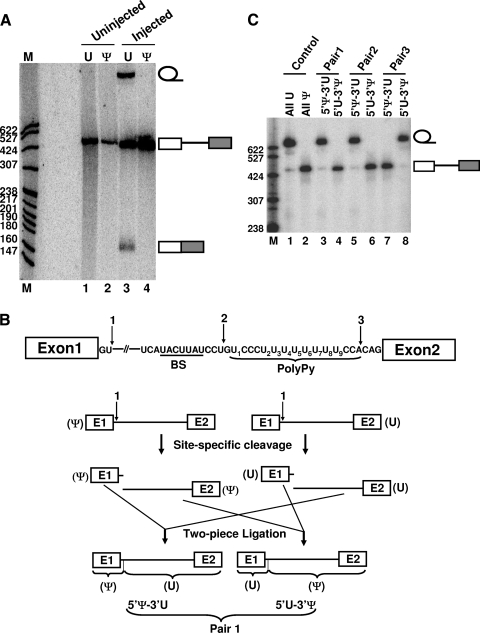
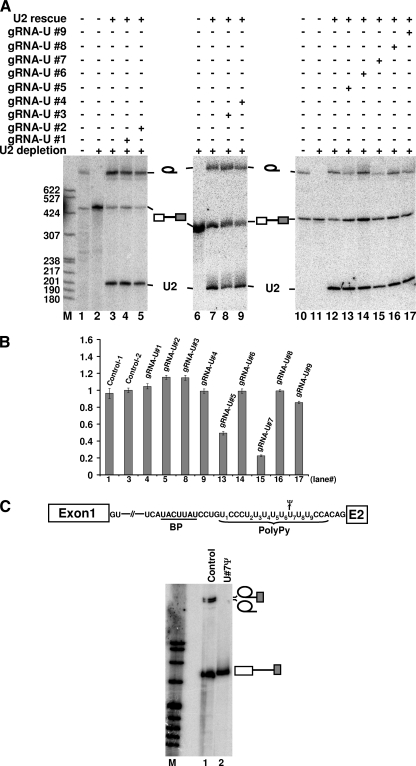
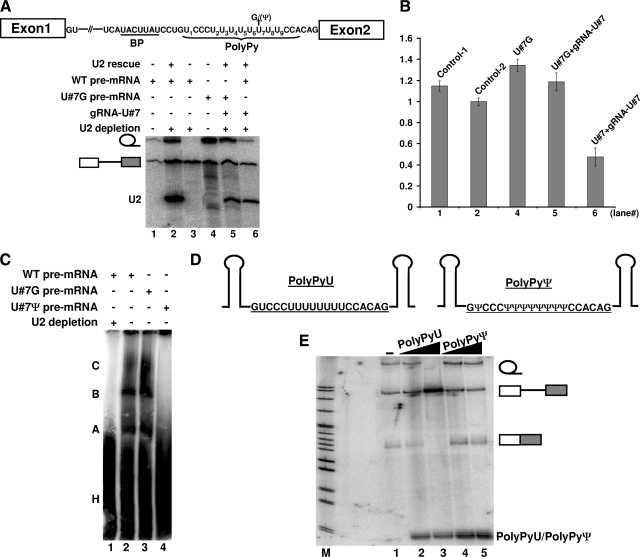
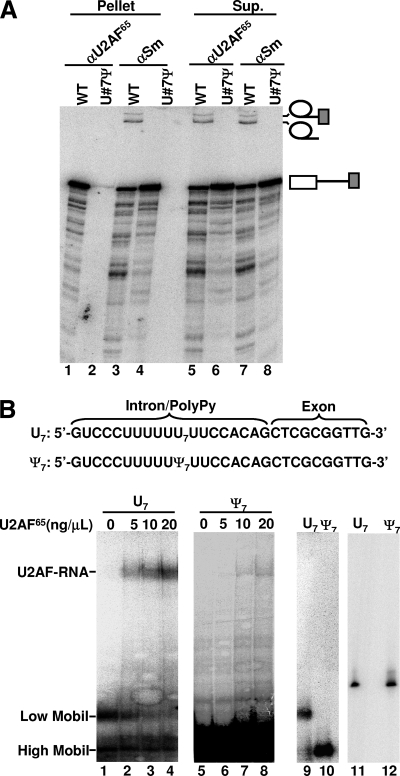
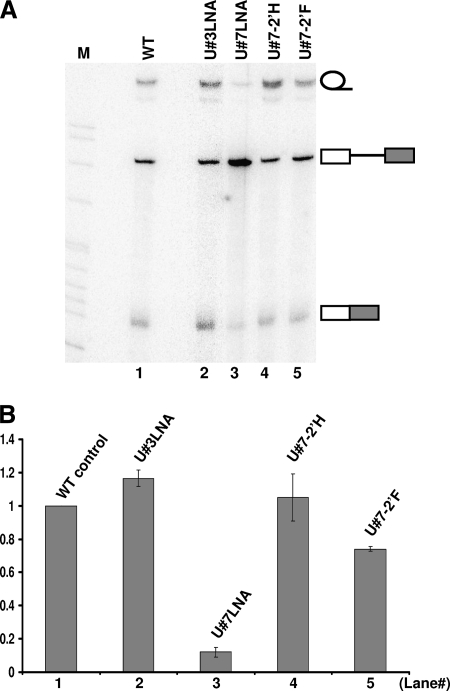
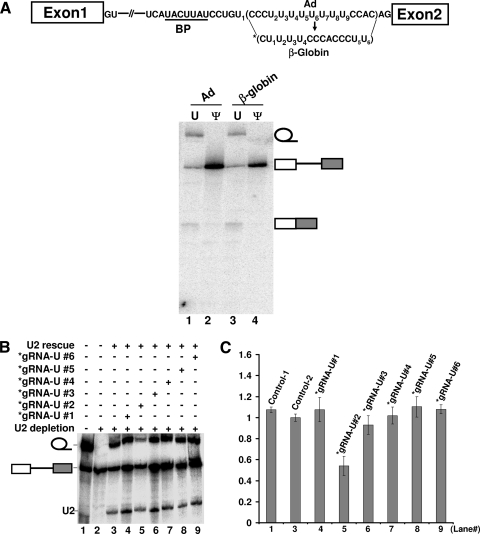
The polypyrimidine tract near the 3' splice site is important for pre-mRNA splicing. Using pseudouridine incorporation and in vivo RNA-guided RNA pseudouridylation, we have identified two important uridines in the polypyrimidine tract of adenovirus pre-mRNA. Conversion of either uridine into pseudouridine leads to a splicing defect in Xenopus oocytes. Using a variety of molecular biology methodologies, we show that the splicing defect is due to the failure of U2AF(65) to recognize the pseudouridylated polypyrimidine tract. This negative impact on splicing is pseudouridine specific, as no effect is observed when the uridine is changed to other naturally occurring nucleotides. Given that pseudouridine favors a C-3'-endo structure, our results suggest that it is backbone flexibility that is key to U2AF binding. Indeed, locking the key uridine in the C-3'-endo configuration while maintaining its uridine identity blocks U2AF(65) binding and splicing. This pseudouridine effect can also be applied to other pre-mRNA polypyrimidine tracts. Thus, our work demonstrates that in vivo binding of U2AF(65) to a polypyrimidine tract requires a flexible RNA backbone.
Figures
References
-
- Adams, M. D., D. Z. Rudner, and D. C. Rio. 1996. Biochemistry and regulation of pre-mRNA splicing. Curr. Opin. Cell Biol. 8:331-839. - PubMed
-
- Arnez, J. G., and T. A. Steitz. 1994. Crystal structure of unmodified tRNA(Gln) complexed with glutaminyl-tRNA synthetase and ATP suggests a possible role for pseudo-uridines in stabilization of RNA structure. Biochemistry 33:7560-7567. - PubMed
-
- Bennett, M., S. Michaud, J. Kingston, and R. Reed. 1992. Protein components specifically associated with prespliceosome and spliceosome complexes. Genes Dev. 6:1986-2000. - PubMed
-
- Burge, C. B., T. Tuschl, and P. A. Sharp. 1999. Splicing of precursors to mRNAs by the spliceosome, p. 525-560. In R. F. Gesteland, T. R. Cech, and J. F. Atkins (ed.), The RNA world, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous