

Review
doi: 10.1007/s11120-010-9577-1.
Epub 2010 Jul 7.
The paleobiological record of photosynthesis
Affiliations
- PMID: 20607406
- PMCID: PMC3021713
- DOI: 10.1007/s11120-010-9577-1
Item in Clipboard
Review
The paleobiological record of photosynthesis
Photosynth Res.
2011 Jan.
Abstract
Fossil evidence of photosynthesis, documented in Precambrian sediments by microbially laminated stromatolites, cyanobacterial microscopic fossils, and carbon isotopic data consistent with the presence of Rubisco-mediated CO2-fixation, extends from the present to ~3,500 million years ago. Such data, however, do not resolve time of origin of O2-producing photoautotrophy from its anoxygenic, bacterial, evolutionary precursor. Though it is well established that Earth's ecosystem has been based on autotrophy since its very early stages, the time of origin of oxygenic photosynthesis, more than 2,450 million years ago, has yet to be established.
Figures

Modern and fossil stromatolites. a Modern stromatolites at Shark Bay (Hamelin Pool), Western Australia. b Modern Shark Bay columnar and domical stromatolites for comparison with (c) fossil stromatolites from the ~2,300-Ma-old Transvaal Dolomite, Cape Province, South Africa. d–f Modern and fossil vertically sliced columnar to domical stromatolites showing upwardly accreted microbial laminae from Shark Bay (d), the ~1,300-Ma-old Belt Supergroup of Montana (e), and the ~3,350-Ma-old Fig Tree Group of the eastern Transvaal, South Africa (f). Scale for a and c shown by the geological hammers enclosed by red circles
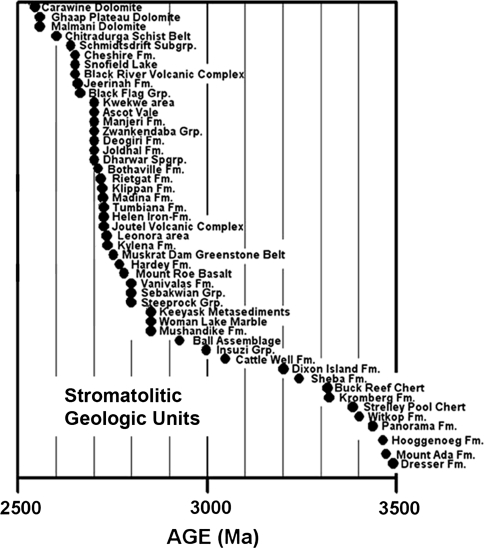
Forty-eight Archean geological units reported to contain stromatolites. Data from Hofmann (2000) and Schopf (2006)

Archean-age microbially laminated stromatolites. a Domical, pseudocolumnar and branching stromatolites, overlain by rippled sediments, and b a domical stromatolite from the ~2,723-Ma-old Tumbiana Formation (Fortescue Group) of Western Australia. c Conical stromatolite and d stratiform and conical stromatolites, from the ~2,985-Ma-old Insuzi Group, South Africa. e–g Laterally linked conical stromatolites from the ~3,388-Ma-old Strelley Pool Chert of Western Australia
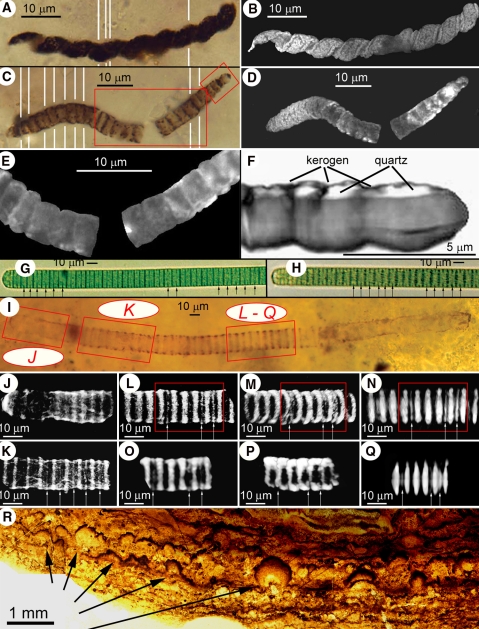
Fossil oscillatoriacean cyanobacteria (a through f) in petrographic thin sections of stromatolitic chert from the ~850-Ma-old Bitter Springs Formation of central Australia; modern oscillatoriaceans (g and h) compared with a morphologically similar fossil trichome (i through q) in a thin section of a cherty stromatolite from the ~775 Ma-old Chichkan Formation of southern Kazakhstan; and pustular laminae, formed by colonies of entophysalidacean cyanobacteria, in a thin section of stromatolitic chert from the ~2,100-Ma-old Kasegalik Formation of the Belcher Islands, Canada. a, b Optical montage (a), composed of five photomicrographs (denoted by the white lines), and a CLSM image (b) of Heliconema, a spirally coiled oscillatoriacean similar to modern Spirulina. c–f Optical montage (c), composed of ten photomicrographs (denoted by the white lines), and CLSM (d and e) and a 3-D Raman image (f) of a large-celled specimen of Cephalophytarion that descends from where it transects the upper surface of the thin section (at the far right) to a depth of 20 μm (at the far left); the larger red rectangle in c denotes the area shown in e, whereas the smaller rectangle denotes the area shown in f; unlike the composite optical image (c), which shows only the medial plane of the specimen, the CLSM image (d) shows its true 3-D morphology; the 3-D Raman image of its end cells (f), rotated to show the flat uppermost surface of the cells where they transect the thin section surface, demonstrates that the kerogenous cell walls (gray) enclose quartz-filled cell lumina (white). g, h Optical images of two specimens of modern Oscillatoria sp. showing the rounded terminal cells (left), disk-shaped medial cells, and partial septations (arrows) characteristic of oscillatoriacean cyanobacteria. i Optical image of the fossil oscillatoriacean, Oscillatoriopsis media, descending into a thin section at a low angle from left to right, shown in a photomontage in which the red rectangles denote the areas of the trichome shown in CLSM images (j through n) and 3-D Raman images (o through q). j The trichome terminus, showing its rounded end-cell and subtending disk-shaped medial cells. k A part of the trichome situated ~14 μm deeper in the section than the trichome terminus (and ~28 μm below the upper surface of the section) that exhibits partial septations (arrows) like those shown in g and h. l–n A deeper part of the trichome (~39 μm below the upper surface of the section) that similarly exhibits partial septations (arrows), in l and m showing the specimen as viewed from above its upper surface (the same perspective as shown in i, but in m with the trichome tilted slightly to the right to show its interior) and in n showing the trichome as viewed from its side. o–q 3-D Raman images (acquired in a spectral window centered in the kerogen “G” band at ~1605 cm−1) showing the kerogenous composition of the trichome and its partial septations: o, the part of specimen denoted by the red rectangle in l, as viewed from above the trichome; p, the part denoted in m, titled slightly to the left; q, the part denoted in n, showing the specimen from its side. r A low-magnification optical image of stromatolitic laminae formed by laterally interlinked colonies (at arrows) of the entophysalidacean cyanobacterium Eoentophysalis
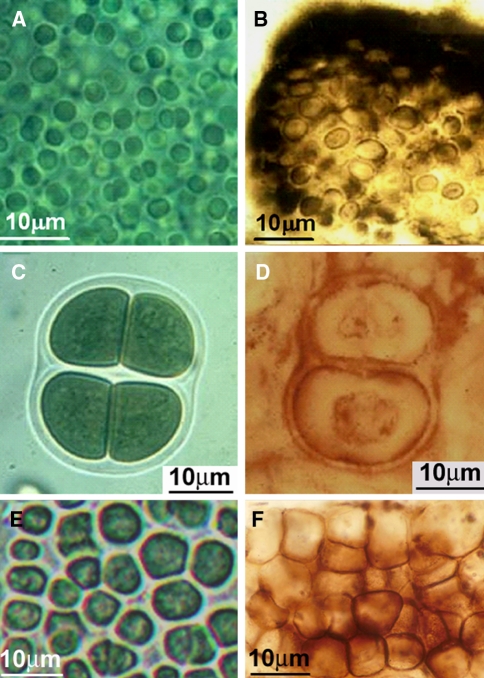
Modern and fossil entophysalidacean, chroococcacean, and pleurocapsacean coccoidal and ellipsoidal cyanobacteria; all fossils are shown in petrographic thin sections of stromatolitic chert. a Modern Entophysalis sp. (Entophysalidaceae) for comparison with b
Eoentophysalis belcherenisis from the ~2,100-Ma-old Kasegalik Formation of the Belcher Islands, Canada. c Modern Gloeocapsa sp. (Chroococcaceae) for comparison with d
Gloeodiniopsis uralicus from the ~1,500-Ma-old Satka Formation of Baskiria, Russia. e Modern Pleurocapsa sp. (PCC 7327, Pleurocapsaceae) for comparison with f
Paleopleurocapsa reniforma from the ~775-Ma-old Chichkan Formation of southern Kazakhstan
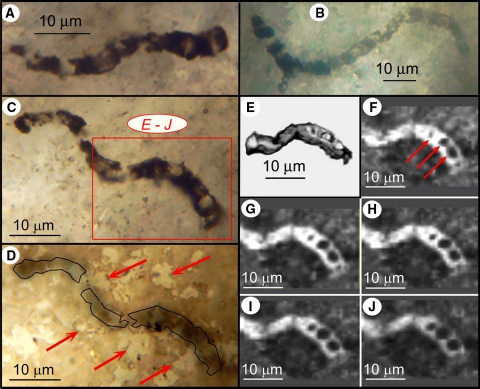
Thin section-embedded filamentous microbes from the ~3,465-Ma-old Apex chert of northwestern Western Australia. a–d Optical images of three specimens of Primaevifilum amoenum, in c and d showing two views of the same specimen situated 3–9 μm below the thin section surface; the red rectangle in c denotes the part shown in e through j; the arrows in d, an optical image of the surface of the thin section with the specimen outlined in black, point to the variously shaped quartz grains of the embedding chert matrix, the irregularity of which shows that the specimen is not a pseudofossil produced by the organic coating of mineral grains. e 3-D Raman image; the organic (carbonaceous, kerogenous) filament (gray) is cylindrical and, like younger Precambrian cellular fossils (e.g., Fig. 3
q), is composed of quartz-filled cells (white). f–j 2-D Raman images at sequential depths below the filament surface (f, at 0.75 μm; g, 1.5 μm; h, 2.25 μm; i, 3.0 μm; j, 3.75 μm); arrows in f point to quartz-filled cell lumina (black) defined by kerogenous cell walls (white), evident also in g through j
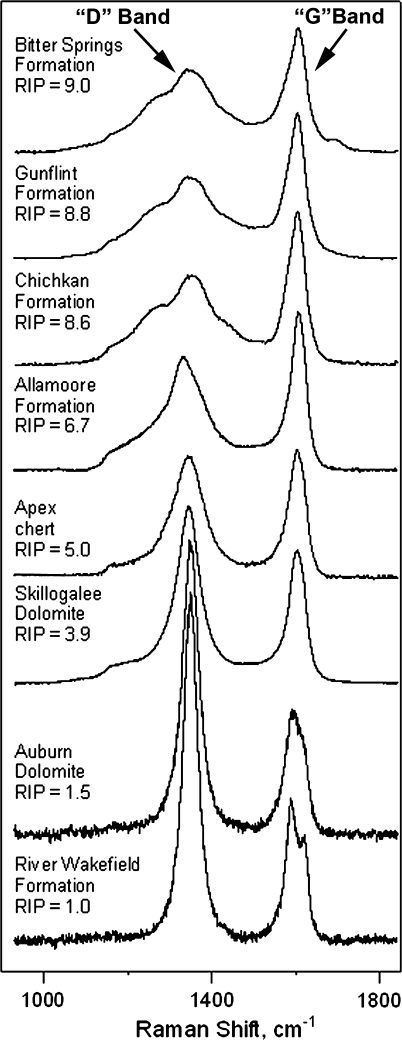
Raman spectra of the kerogenous cell walls of representative Precambrian microfossils permineralized in cherts of the ~850-Ma-old Bitter Springs, ~1900-Ma-old Gunflint, ~775 Ma-old Chichkan, and ~1050-Ma-old Allamoore Formations, the ~3,465-Ma-old Apex chert, the ~760-Ma-old Skillogalee and ~720-Ma-old Auburn Dolomites, and the ~775-Ma-old River Wakefield Formation (Schopf et al. 2005, 2007), ordered by their RIP values (Schopf et al. 2005) from less (top) to more (bottom) geochemically mature
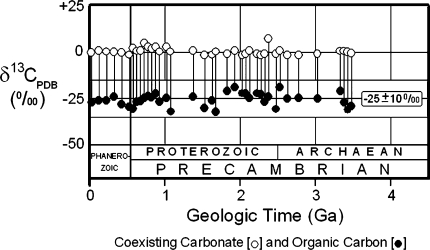
Carbon isotopic values of coexisting carbonate and organic carbon measured in bulk samples of Phanerozoic and Precambrian sedimentary rocks, for the Precambrian represented by data from 100 fossiliferous cherts and shales shown as average values for groups of samples from 50-Ma-long intervals (Strauss and Moore ; Schopf 1994b)
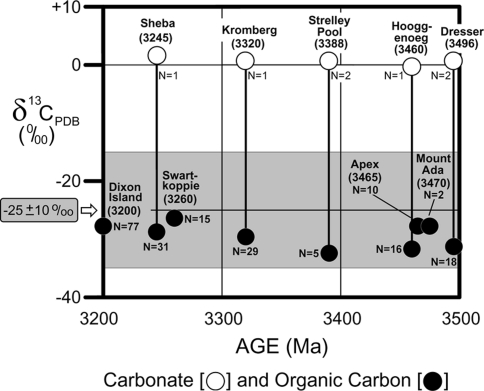
Carbon isotopic values of carbonate and organic carbon measured in bulk samples of the oldest microfossiliferous units now known (Schopf 2006)
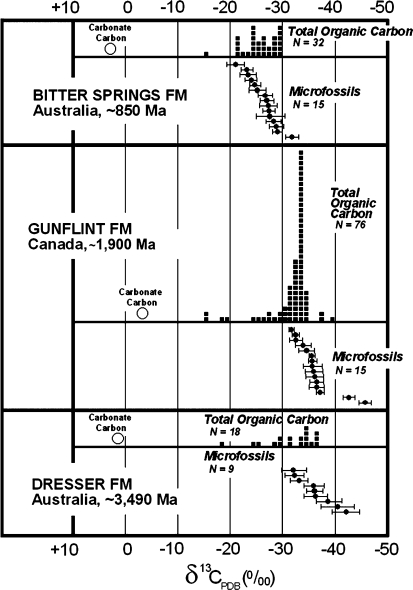
Carbon isotopic values of individual Precambrian microfossils measured by secondary ion microprobe spectrometry (SIMS) compared with those of the carbonate and total organic carbon measured in bulk samples of the same geological units. Values plotted for carbonate and total organic carbon are from Strauss and Moore (1992); for microfossils from the Bitter Springs and Gunflint Formations, from House et al. (2000); and those for microfossils from the Dresser Formation, from Ueno et al. (2001a)
Similar articles
-
Geological evidence of oxygenic photosynthesis and the biotic response to the 2400-2200 ma "great oxidation event".Biochemistry (Mosc). 2014 Mar;79(3):165-77. doi: 10.1134/S0006297914030018. Biochemistry (Mosc). 2014. PMID: 24821442 Review.
-
The role of biology in planetary evolution: cyanobacterial primary production in low-oxygen Proterozoic oceans.Environ Microbiol. 2016 Feb;18(2):325-40. doi: 10.1111/1462-2920.13118. Epub 2015 Dec 21. Environ Microbiol. 2016. PMID: 26549614 Free PMC article. Review.
-
Morphological record of oxygenic photosynthesis in conical stromatolites.Proc Natl Acad Sci U S A. 2009 Jul 7;106(27):10939-43. doi: 10.1073/pnas.0900885106. Epub 2009 Jun 29. Proc Natl Acad Sci U S A. 2009. PMID: 19564621 Free PMC article.
-
The initiation of biological processes on Earth: summary of empirical evidence.Adv Space Res. 1992;12(4):143-56. doi: 10.1016/0273-1177(92)90168-w. Adv Space Res. 1992. PMID: 11538134 Review.
-
Earth's early biosphere.Gravit Space Biol Bull. 1998 May;11(2):23-30. Gravit Space Biol Bull. 1998. PMID: 11540635 Review.
Cited by
-
Timing the Evolutionary Advent of Cyanobacteria and the Later Great Oxidation Event Using Gene Phylogenies of a Sunscreen.mBio. 2019 May 21;10(3):e00561-19. doi: 10.1128/mBio.00561-19. mBio. 2019. PMID: 31113897 Free PMC article.
-
The Origin of RNA and the Formose-Ribose-RNA Pathway.Int J Mol Sci. 2024 Jun 19;25(12):6727. doi: 10.3390/ijms25126727. Int J Mol Sci. 2024. PMID: 38928433 Free PMC article. Review.
-
A Comprehensive Study of Cyanobacterial Morphological and Ecological Evolutionary Dynamics through Deep Geologic Time.PLoS One. 2016 Sep 20;11(9):e0162539. doi: 10.1371/journal.pone.0162539. eCollection 2016. PLoS One. 2016. PMID: 27649395 Free PMC article.
-
Thermal-stable proteins of fruit of long-living Sacred Lotus Nelumbo nucifera Gaertn var. China Antique.Trop Plant Biol. 2013 Sep 1;6(2-3):10.1007/s12042-013-9124-2. doi: 10.1007/s12042-013-9124-2. Trop Plant Biol. 2013. PMID: 24363819 Free PMC article.
-
Cyanotoxins and the Nervous System.Toxins (Basel). 2021 Sep 16;13(9):660. doi: 10.3390/toxins13090660. Toxins (Basel). 2021. PMID: 34564664 Free PMC article. Review.
References
-
- Allwood AC, Kamber BS, Walter MR, Burch IW, Kanik I. Trace elements record depositional history of an Early Archean stromatolitic carbonate platform. Chem Geol. 2010;270:148–163. doi: 10.1016/j.chemgeo.2009.11.013. - DOI
