

Role of WASP in cell polarity and podosome dynamics of myeloid cells
- PMID: 20609498
- PMCID: PMC3037472
- DOI: 10.1016/j.ejcb.2010.05.009
Role of WASP in cell polarity and podosome dynamics of myeloid cells
Abstract
The integrin-dependent migration of myeloid cells requires tight coordination between actin-based cell membrane protrusion and integrin-mediated adhesion to form a stable leading edge. Under this mode of migration, polarised myeloid cells including dendritic cells, macrophages and osteoclasts develop podosomes that sustain the extending leading edge. Podosome integrity and dynamics vary in response to changes in the physical and biochemical properties of the cell environment. In the current article we discuss the role of various factors in initiation and stability of podosomes and the roles of the Wiskott Aldrich Syndrome Protein (WASP) in this process. We discuss recent data indicating that in a cellular context WASP is crucial not only for localised actin polymerisation at the leading edge and in podosome cores but also for coordination of integrin clustering and activation during podosome formation and disassembly.
Copyright © 2010 Elsevier GmbH. All rights reserved.
Figures
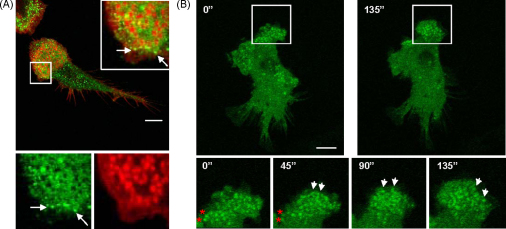
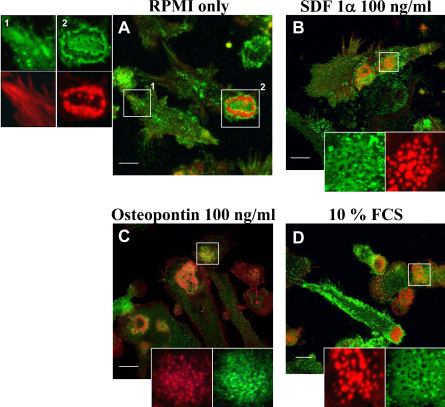
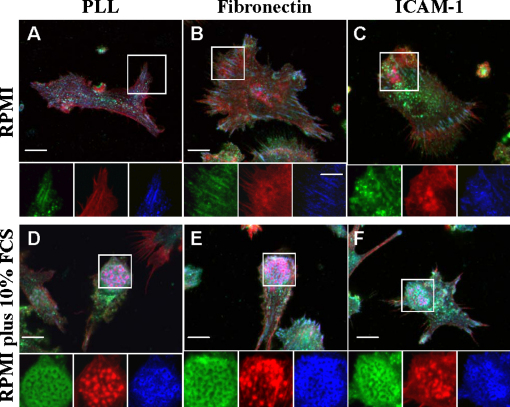
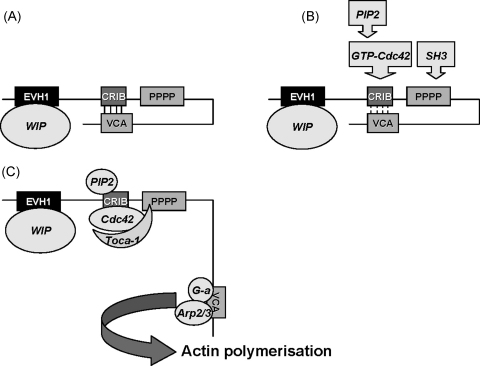
Similar articles
-
Tyrosine phosphorylation of WASP promotes calpain-mediated podosome disassembly.Haematologica. 2012 May;97(5):687-91. doi: 10.3324/haematol.2011.048868. Epub 2011 Dec 1. Haematologica. 2012. PMID: 22133775 Free PMC article.
-
Integrin linked kinase (ILK) regulates podosome maturation and stability in dendritic cells.Int J Biochem Cell Biol. 2014 May;50(100):47-54. doi: 10.1016/j.biocel.2014.01.021. Epub 2014 Feb 6. Int J Biochem Cell Biol. 2014. PMID: 24508783 Free PMC article.
-
Wiskott-Aldrich syndrome protein and the cytoskeletal dynamics of dendritic cells.J Pathol. 2004 Nov;204(4):460-9. doi: 10.1002/path.1651. J Pathol. 2004. PMID: 15495215 Review.
-
Hematopoietic lineage cell-specific protein 1 functions in concert with the Wiskott-Aldrich syndrome protein to promote podosome array organization and chemotaxis in dendritic cells.J Immunol. 2011 Apr 15;186(8):4805-18. doi: 10.4049/jimmunol.1003102. Epub 2011 Mar 11. J Immunol. 2011. PMID: 21398607 Free PMC article.
-
WASP-interacting protein (WIP): working in polymerisation and much more.Trends Cell Biol. 2007 Nov;17(11):555-62. doi: 10.1016/j.tcb.2007.08.005. Epub 2007 Oct 18. Trends Cell Biol. 2007. PMID: 17949983 Review.
Cited by
-
Cdc42 regulates neutrophil migration via crosstalk between WASp, CD11b, and microtubules.Blood. 2012 Oct 25;120(17):3563-74. doi: 10.1182/blood-2012-04-426981. Epub 2012 Aug 29. Blood. 2012. PMID: 22932798 Free PMC article.
-
Costameres, dense plaques and podosomes: the cell matrix adhesions in cardiovascular mechanosensing.J Muscle Res Cell Motil. 2019 Jun;40(2):197-209. doi: 10.1007/s10974-019-09529-7. Epub 2019 Jun 18. J Muscle Res Cell Motil. 2019. PMID: 31214894 Free PMC article. Review.
-
Selenium nanoparticles induce suppressed function of tumor associated macrophages and inhibit Dalton's lymphoma proliferation.Biochem Biophys Rep. 2017 Sep 21;12:172-184. doi: 10.1016/j.bbrep.2017.09.005. eCollection 2017 Dec. Biochem Biophys Rep. 2017. PMID: 29090279 Free PMC article.
-
Bayesian localization microscopy reveals nanoscale podosome dynamics.Nat Methods. 2011 Dec 4;9(2):195-200. doi: 10.1038/nmeth.1812. Nat Methods. 2011. PMID: 22138825 Free PMC article.
-
Wiskott-Aldrich syndrome protein-deficient hematopoietic cells can be efficiently mobilized by granulocyte colony-stimulating factor.Haematologica. 2013 Aug;98(8):1300-8. doi: 10.3324/haematol.2012.077040. Epub 2013 Feb 26. Haematologica. 2013. PMID: 23445877 Free PMC article.
References
-
- Ancliff P.J., Blundell M.P., Cory G.O., Calle Y., Worth A., Kempski H., Burns S., Jones G.E., Sinclair J., Kinnon C., Hann I.M., Gale R.E., Linch D.C., Thrasher A.J. Two novel activating mutations in the Wiskott–Aldrich syndrome protein result in congenital neutropenia. Blood. 2006;108:2182–2189. - PubMed
-
- Anton I.M., Jones G.E. WIP: a multifunctional protein involved in actin cytoskeleton regulation. Eur. J. Cell. Biol. 2006;85:295–304. - PubMed
-
- Anton I.M., Jones G.E., Wandosell F., Geha R., Ramesh N. WASP-interacting protein (WIP): working in polymerisation and much more. Trends Cell. Biol. 2007;17:555–562. - PubMed
-
- Blundell M.P., Bouma G., Calle Y., Jones G.E., Kinnon C., Thrasher A.J. Improvement of migratory defects in a murine model of Wiskott–Aldrich syndrome gene therapy. Mol. Ther. 2008;16:836–844. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
