

A random cell motility gradient downstream of FGF controls elongation of an amniote embryo
- PMID: 20613841
- PMCID: PMC3118990
- DOI: 10.1038/nature09151
A random cell motility gradient downstream of FGF controls elongation of an amniote embryo
Abstract
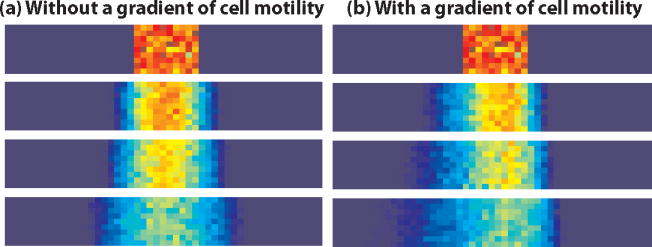
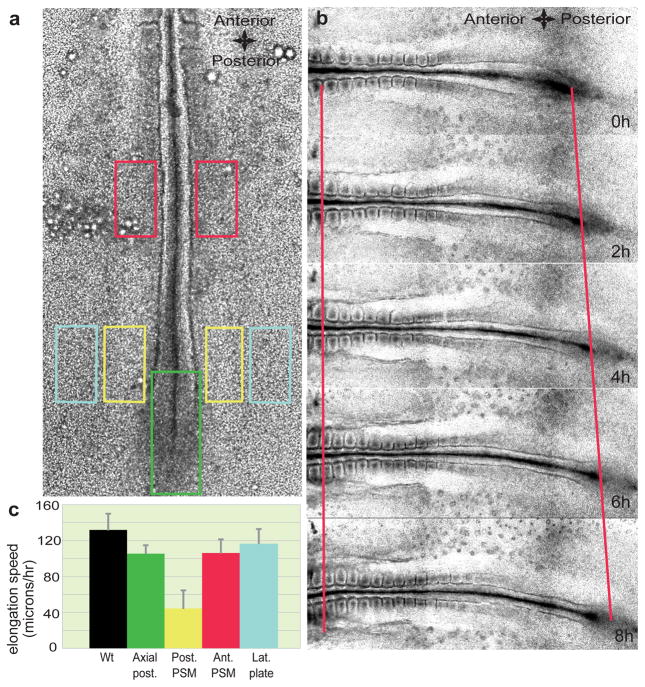
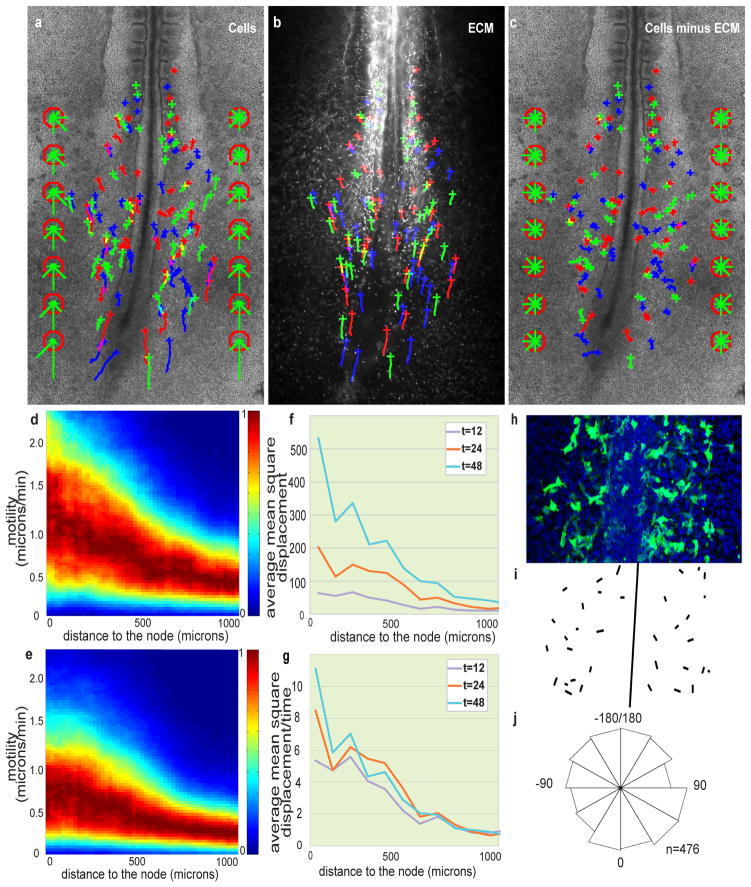
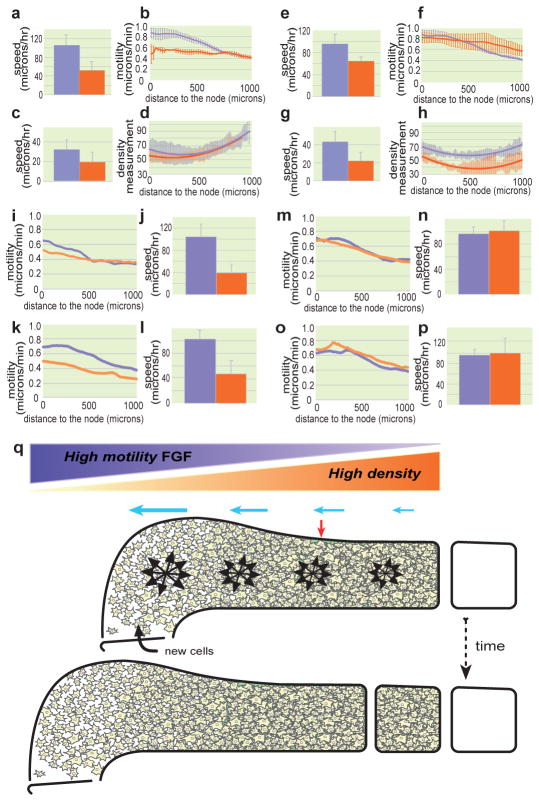
Vertebrate embryos are characterized by an elongated antero-posterior (AP) body axis, which forms by progressive cell deposition from a posterior growth zone in the embryo. Here, we used tissue ablation in the chicken embryo to demonstrate that the caudal presomitic mesoderm (PSM) has a key role in axis elongation. Using time-lapse microscopy, we analysed the movements of fluorescently labelled cells in the PSM during embryo elongation, which revealed a clear posterior-to-anterior gradient of cell motility and directionality in the PSM. We tracked the movement of the PSM extracellular matrix in parallel with the labelled cells and subtracted the extracellular matrix movement from the global motion of cells. After subtraction, cell motility remained graded but lacked directionality, indicating that the posterior cell movements associated with axis elongation in the PSM are not intrinsic but reflect tissue deformation. The gradient of cell motion along the PSM parallels the fibroblast growth factor (FGF)/mitogen-activated protein kinase (MAPK) gradient, which has been implicated in the control of cell motility in this tissue. Both FGF signalling gain- and loss-of-function experiments lead to disruption of the motility gradient and a slowing down of axis elongation. Furthermore, embryos treated with cell movement inhibitors (blebbistatin or RhoK inhibitor), but not cell cycle inhibitors, show a slower axis elongation rate. We propose that the gradient of random cell motility downstream of FGF signalling in the PSM controls posterior elongation in the amniote embryo. Our data indicate that tissue elongation is an emergent property that arises from the collective regulation of graded, random cell motion rather than by the regulation of directionality of individual cellular movements.
Figures
References
-
- Dubrulle J, Pourquie O. fgf8 mRNA decay establishes a gradient that couples axial elongation to patterning in the vertebrate embryo. Nature. 2004;427:419–422. - PubMed
-
- Keller R, Shook D, Skoglund P. The forces that shape embryos: physical aspects of convergent extension by cell intercalation. Physical biology. 2008;5:15007. - PubMed
-
- Lawson A, Schoenwolf GC. New insights into critical events of avian gastrulation. Anat Rec. 2001;262:238–252. - PubMed
-
- Voiculescu O, Bertocchini F, Wolpert L, Keller RE, Stern CD. The amniote primitive streak is defined by epithelial cell intercalation before gastrulation. Nature. 2007;449:1049–1052. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
