

Conformational selection in the molten globule state of the nuclear coactivator binding domain of CBP
- PMID: 20616042
- PMCID: PMC2906600
- DOI: 10.1073/pnas.1001693107
Conformational selection in the molten globule state of the nuclear coactivator binding domain of CBP
Abstract
Native molten globules are the most folded kind of intrinsically disordered proteins. Little is known about the mechanism by which native molten globules bind to their cognate ligands to form fully folded complexes. The nuclear coactivator binding domain (NCBD) of CREB binding protein is particularly interesting in this respect as structural studies of its complexes have shown that NCBD folds into two remarkably different states depending on the ligand being ACTR or IRF-3. The ligand-free state of NCBD was characterized in order to understand the mechanism of folding upon ligand binding. Biophysical studies show that despite the molten globule nature of the domain, it contains a small cooperatively folded core. By NMR spectroscopy, we have demonstrated that the folded core of NCBD has a well ordered conformer with specific side chain packing. This conformer resembles the structure of the NCBD in complex with the protein ligand, ACTR, suggesting that ACTR binds to prefolded NCBD molecules from the ensemble of interconverting structures.
Conflict of interest statement
The authors declare no conflict of interest.
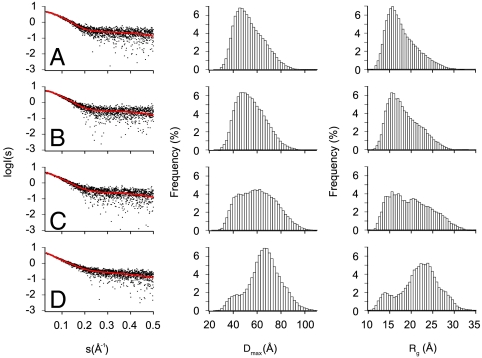
Figures




References
-
- Wright PE, Dyson HJ. Intrinsically unstructured proteins: re-assessing the protein structure-function paradigm. J Mol Biol. 1999;293:321–331. - PubMed
-
- Dunker AK, Brown CJ, Lawson JD, Iakoucheva LM, Obradovic Z. Intrinsic disorder and protein function. Biochemistry. 2002;41:6573–6582. - PubMed
-
- Dyson HJ, Wright PE. Intrinsically unstructured proteins and their functions. Nat Rev Mol Cell Biol. 2005;6:197–208. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
