

Important roles of PI3Kgamma in osteoclastogenesis and bone homeostasis
- PMID: 20616072
- PMCID: PMC2919938
- DOI: 10.1073/pnas.1001499107
Important roles of PI3Kgamma in osteoclastogenesis and bone homeostasis
Abstract
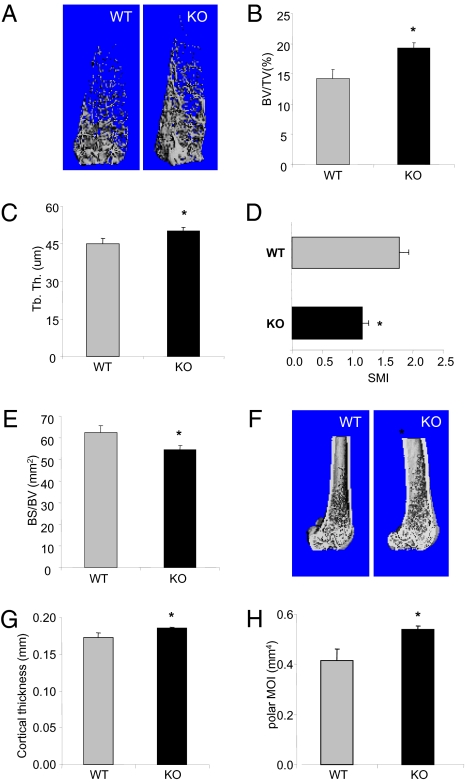
G protein-coupled receptor-regulated PI3Kgamma is abundantly expressed in myeloid cells and has been implicated as a promising drug target to treat various inflammatory diseases. However, its role in bone homeostasis has not been investigated, despite the fact that osteoclasts are derived from myeloid lineage. We therefore carried out thorough bone phenotypic characterization of a PI3Kgamma-deficient mouse line and found that PI3Kgamma-deficient mice had high bone mass. Our analyses further revealed that PI3Kgamma deficiency did not affect bone formation because no significant changes in osteoblast number and bone formation rate were observed. Instead, the lack of PI3Kgamma was associated with decreased bone resorption, as evidenced by decreased osteoclast number in vivo and impaired osteoclast formation in vitro. The decreased osteoclast formation was accompanied by down-regulated expression of osteoclastogenic genes, compromised chemokine receptor signaling, and an increase in apoptosis during osteoclast differentiation. Together, these data suggest that PI3Kgamma regulates bone homeostasis by modulating osteoclastogenesis. Our study also suggests that inhibition of PI3Kgamma, which is being considered as a potential therapeutic strategy for treating chronic inflammatory disorders, may result in an increase in bone mass.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Cantley LC. The phosphoinositide 3-kinase pathway. Science. 2002;296:1655–1657. - PubMed
-
- Hirsch E, et al. Central role for G protein-coupled phosphoinositide 3-kinase gamma in inflammation. Science. 2000;287:1049–1053. - PubMed
-
- Laffargue M, et al. Phosphoinositide 3-kinase gamma is an essential amplifier of mast cell function. Immunity. 2002;16:441–451. - PubMed
-
- Barber DF, et al. PI3Kgamma inhibition blocks glomerulonephritis and extends lifespan in a mouse model of systemic lupus. Nat Med. 2005;11:933–935. - PubMed
-
- Camps M, et al. Blockade of PI3Kgamma suppresses joint inflammation and damage in mouse models of rheumatoid arthritis. Nat Med. 2005;11:936–943. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
