

Lung self-assembly is modulated by tissue surface tensions
- PMID: 20616358
- PMCID: PMC3095985
- DOI: 10.1165/rcmb.2009-0309OC
Lung self-assembly is modulated by tissue surface tensions
Abstract
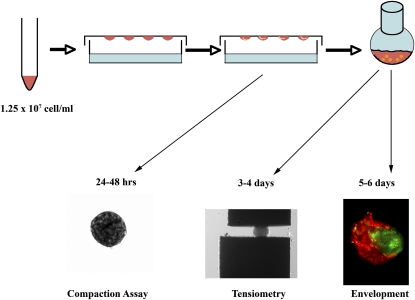
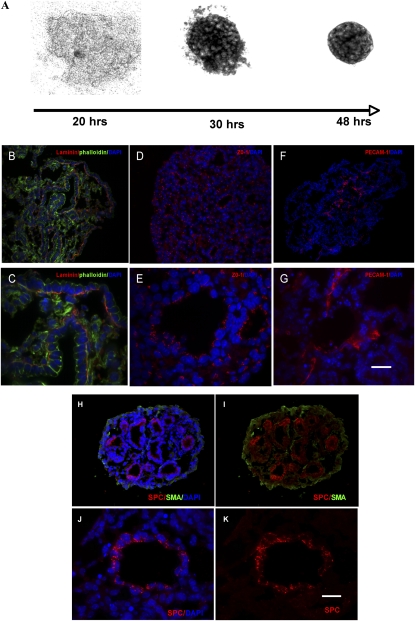
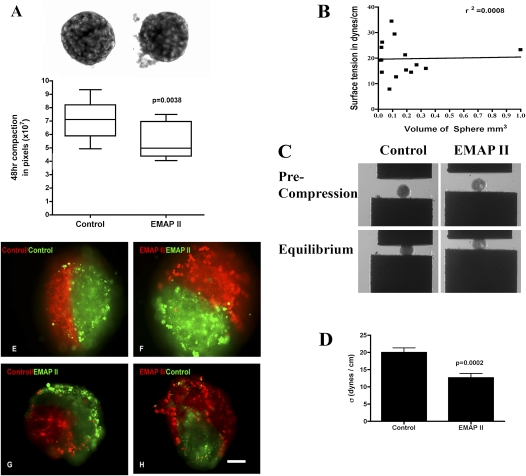
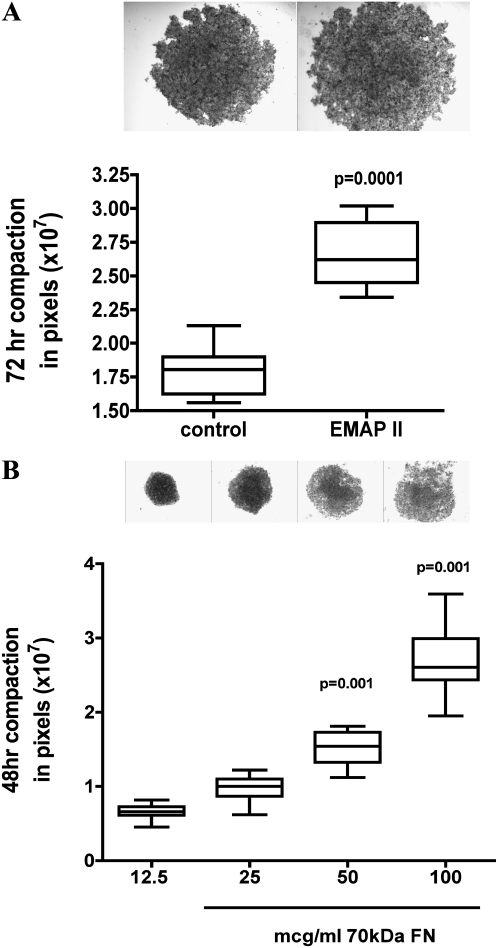
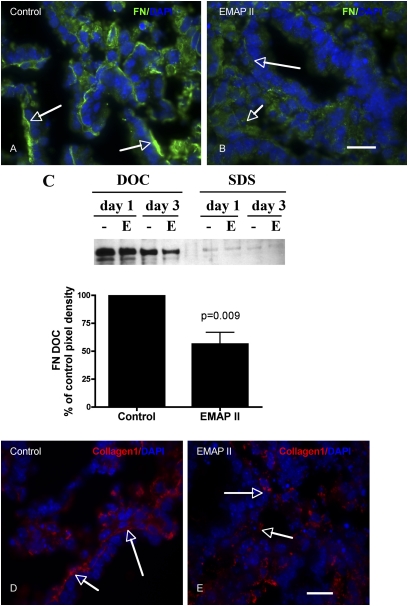
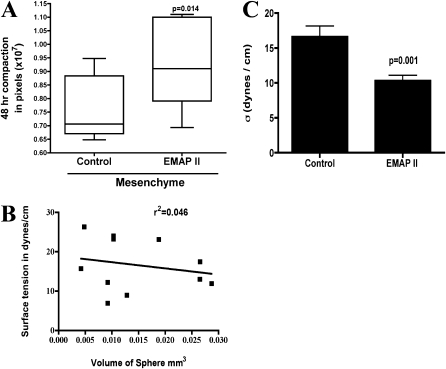
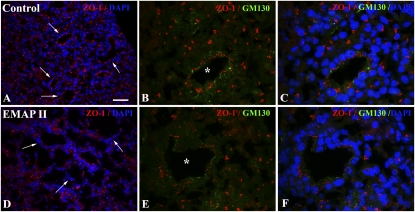
To identify cell-intrinsic properties that facilitate interaction between epithelial endodermal and mesenchymal mesodermal cells during lung morphogenesis, we developed a model of lung self-assembly that mimics fetal lung formation in structure, polarity, vasculature, and extracellular matrix expression. Three-dimensional pulmonary bodies (PBs) spontaneously self-assemble from single-cell suspensions and exhibit liquid-like properties that allow measurements of compaction rate and cohesion, and that may help to specify cellular self-organization. We hypothesized that changes in one or more of these parameters could potentially explain the lung hypoplasia associated with abnormal lung development. We examined the impact of endothelial/monocyte-activating polypeptide (EMAP) II in PBs, because EMAPII is highly expressed in lung hypoplasia. EMAPII significantly increased compaction rate and decreased overall cohesion of PBs composed of both epithelial and mesenchymal cells. Moreover, the effects of EMAPII on compaction and cohesion act exclusively through the mesenchymal cell population by interfering with fibronectin matrix assembly. We also show that EMAPII alters epithelial cell polarity and surfactant protein C expression. Our findings demonstrate, for the first time, that PBs possess liquid-like properties that can help to guide the self-assembly of fetal lungs, and that EMAPII expression can influence both mesenchymal and epithelial cells but through different molecular mechanisms.
Figures
References
-
- Zeng X, Wert SE, Federici R, Peters KG, Whitsett JA. VEGF enhances pulmonary vasculogenesis and disrupts lung morphogenesis in vivo. Dev Dyn 1998;211:215–227. - PubMed
-
- Schwarz MA, Zhang F, Gebb S, Starnes V, Warburton D. EMAP II inhibits lung neovascularization and airway epithelial morphogenesis. Mech Dev 2000;95:123–132. - PubMed
-
- Dickson MC, Martin JS, Cousins FM, Kulkarni AB, Karlsson S, Akhurst RJ. Defective haematopoiesis and vasculogenesis in transforming growth factor–beta 1 knock out mice. Development 1995;121:1845–1854. - PubMed
-
- Zhou L, Dey CR, Wert SE, Whitsett JA. Arrested lung morphogenesis in transgenic mice bearing an SP-C–TGF–beta 1 chimeric gene. Dev Biol 1996;175:227–238. - PubMed
-
- Schuger L, O'Shea KS, Nelson BB, Varani J. Organotypic arrangement of mouse embryonic lung cells on a basement membrane extract: involvement of laminin. Development 1990;110:1091–1099. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
