

Comparative analysis of DNA replication timing reveals conserved large-scale chromosomal architecture
- PMID: 20617169
- PMCID: PMC2895651
- DOI: 10.1371/journal.pgen.1001011
Comparative analysis of DNA replication timing reveals conserved large-scale chromosomal architecture
Abstract
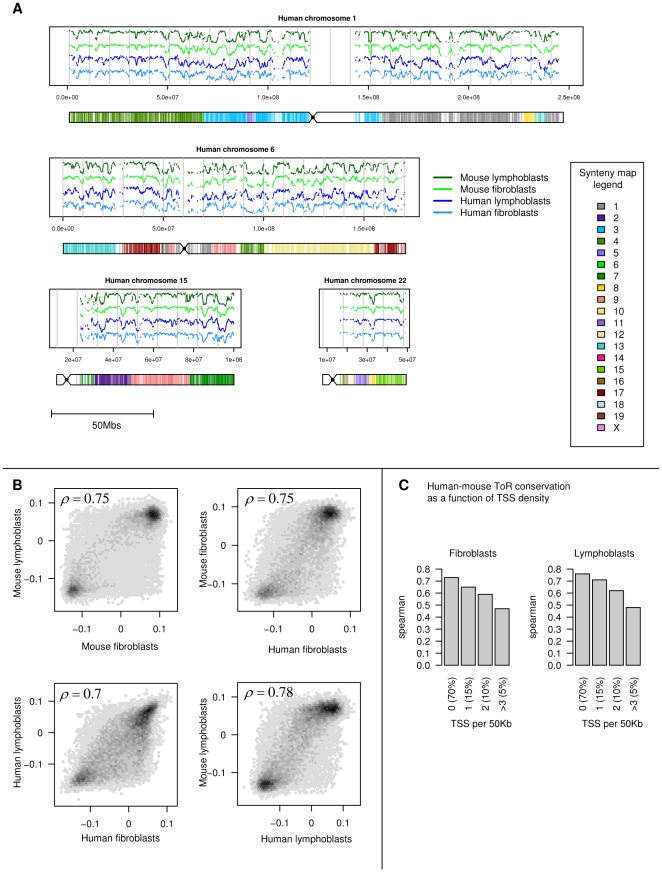
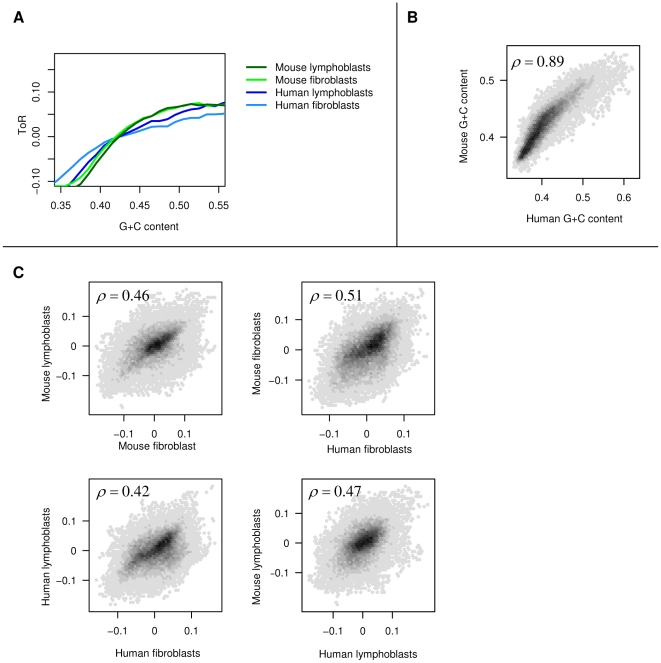
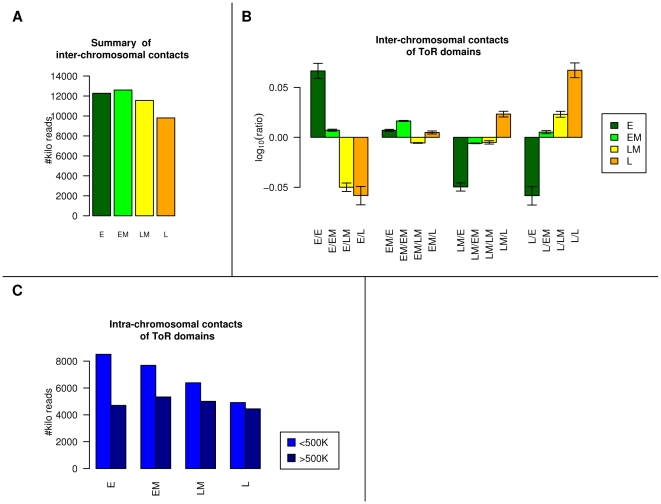
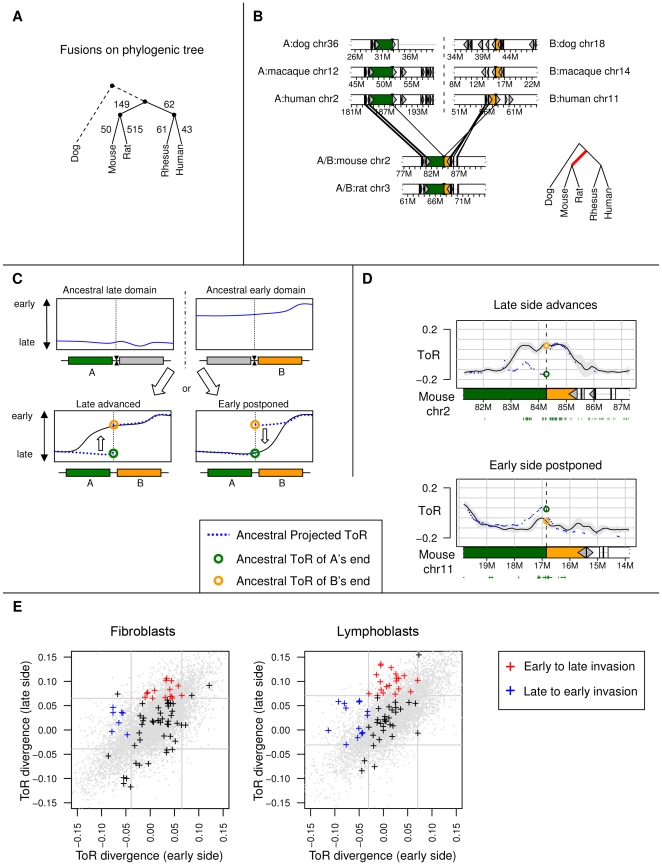
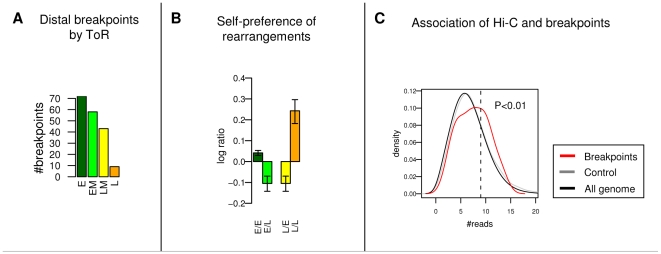
Recent evidence suggests that the timing of DNA replication is coordinated across megabase-scale domains in metazoan genomes, yet the importance of this aspect of genome organization is unclear. Here we show that replication timing is remarkably conserved between human and mouse, uncovering large regions that may have been governed by similar replication dynamics since these species have diverged. This conservation is both tissue-specific and independent of the genomic G+C content conservation. Moreover, we show that time of replication is globally conserved despite numerous large-scale genome rearrangements. We systematically identify rearrangement fusion points and demonstrate that replication time can be locally diverged at these loci. Conversely, rearrangements are shown to be correlated with early replication and physical chromosomal proximity. These results suggest that large chromosomal domains of coordinated replication are shuffled by evolution while conserving the large-scale nuclear architecture of the genome.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Bernstein BE, Mikkelsen TS, Xie X, Kamal M, Huebert DJ, et al. A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell. 2006;125:315–326. - PubMed
-
- Gilbert N, Boyle S, Fiegler H, Woodfine K, Carter NP, et al. Chromatin architecture of the human genome: gene-rich domains are enriched in open chromatin fibers. Cell. 2004;118:555–566. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
