

Hematopoietic stem cell quiescence promotes error-prone DNA repair and mutagenesis
- PMID: 20619762
- PMCID: PMC2924905
- DOI: 10.1016/j.stem.2010.06.014
Hematopoietic stem cell quiescence promotes error-prone DNA repair and mutagenesis
Abstract
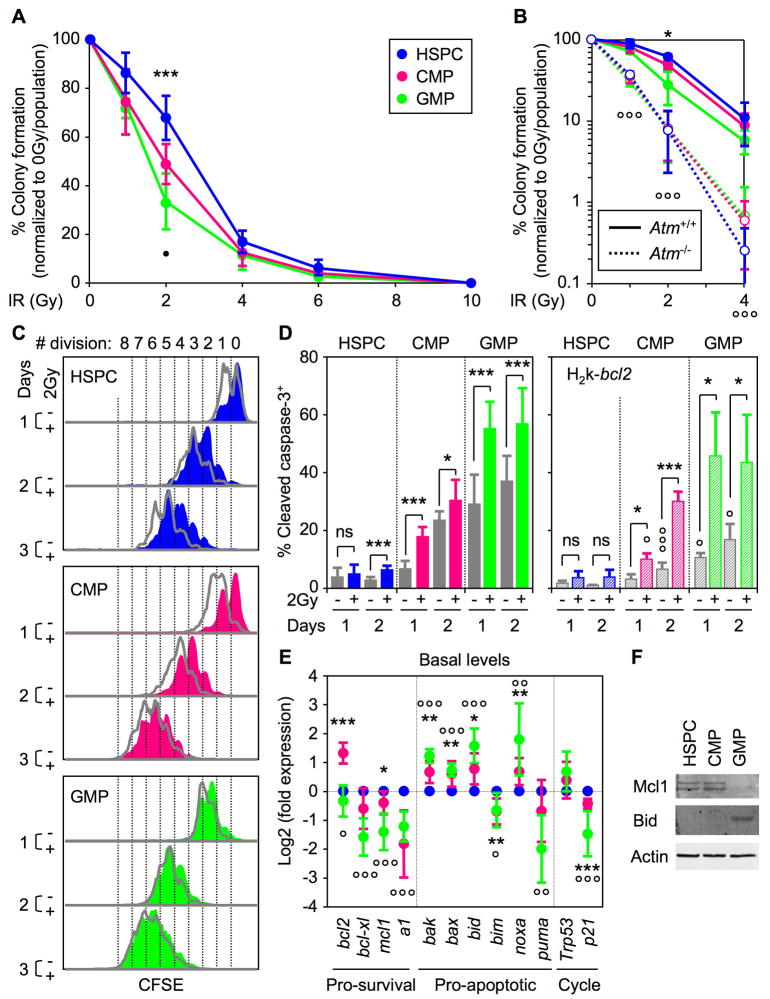
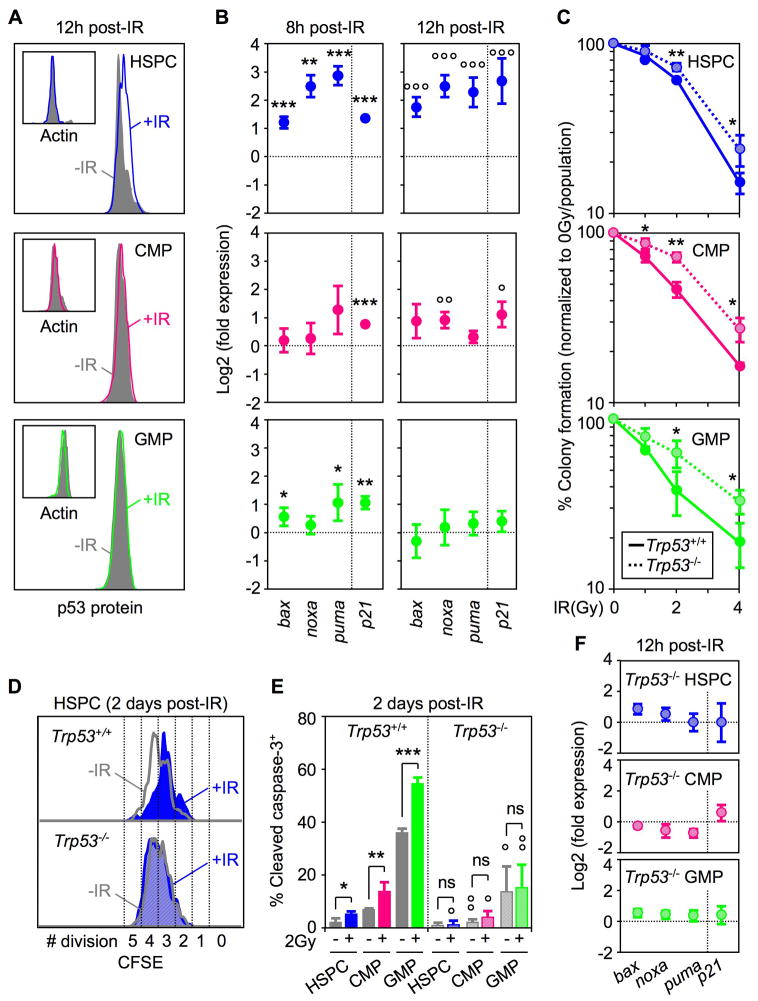
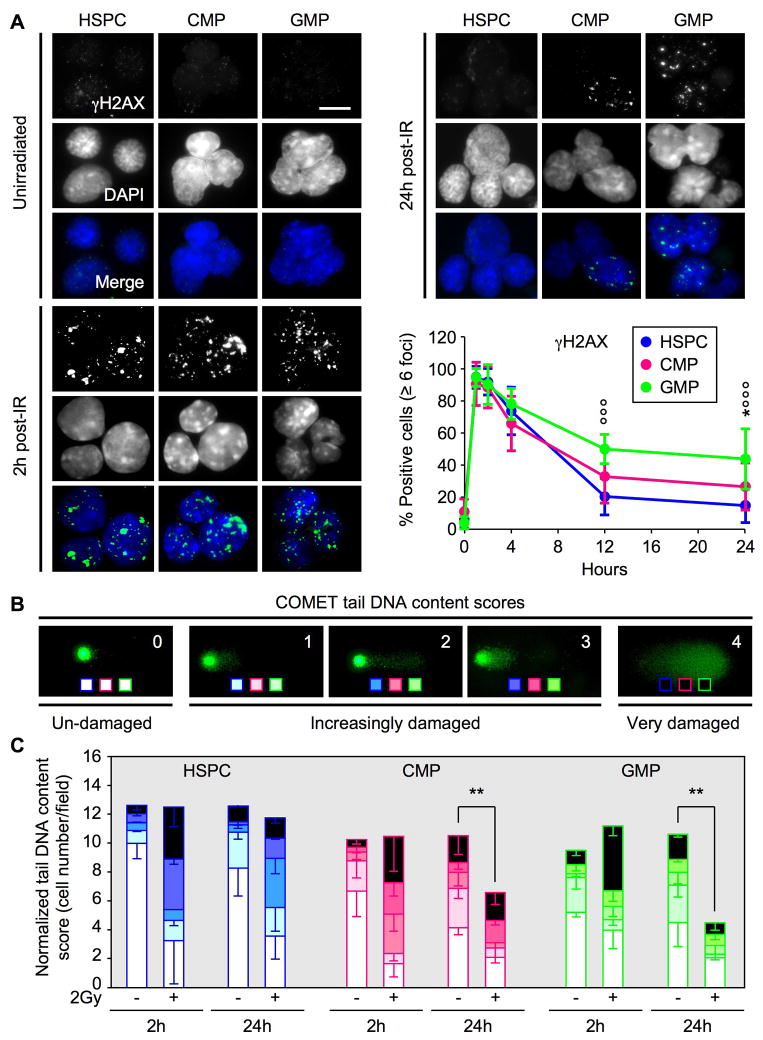
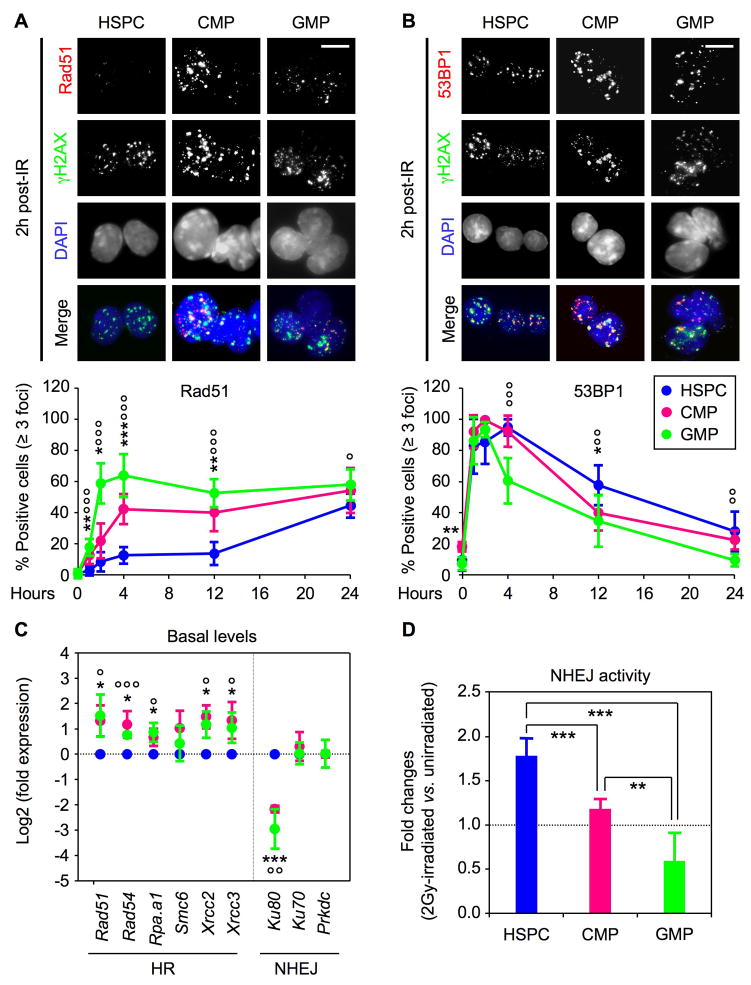
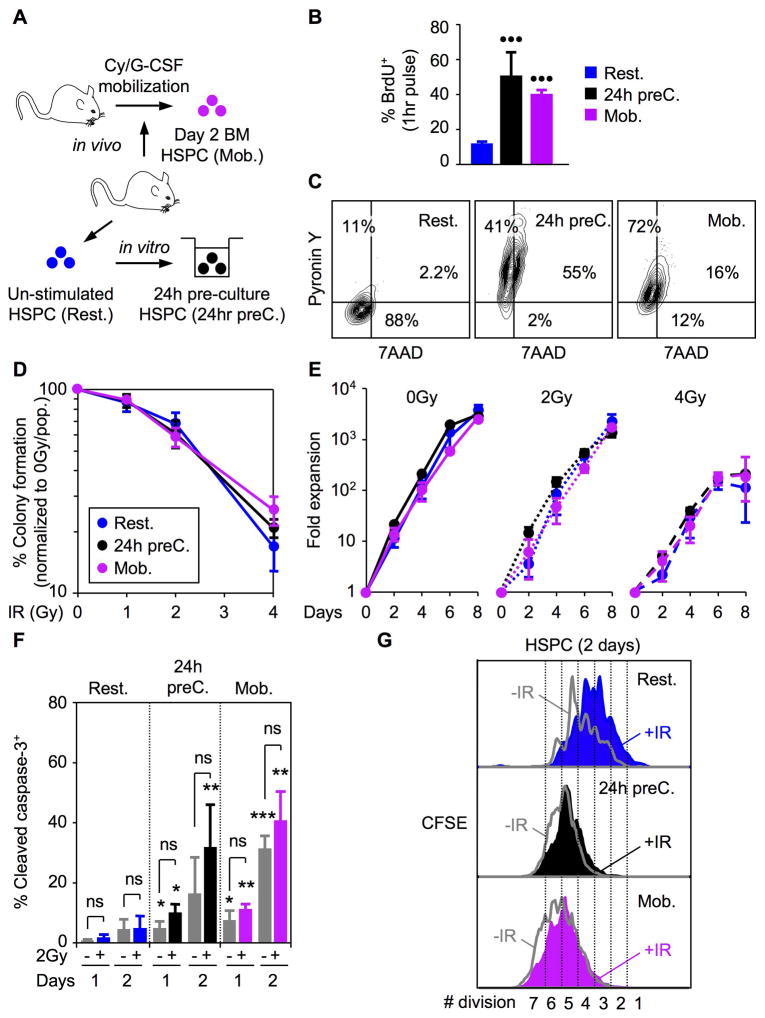
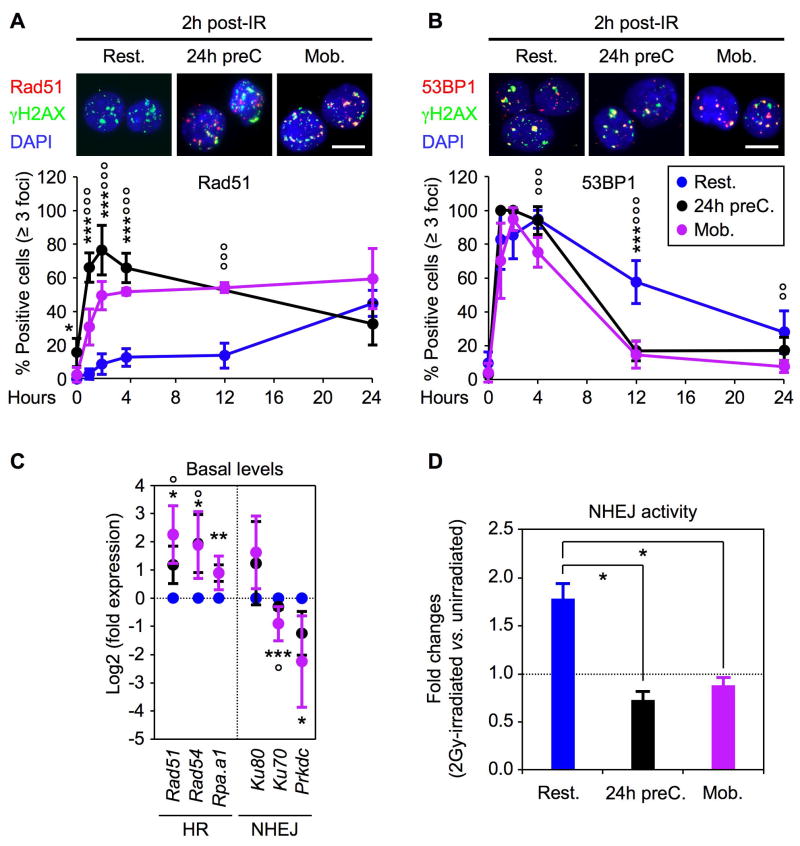
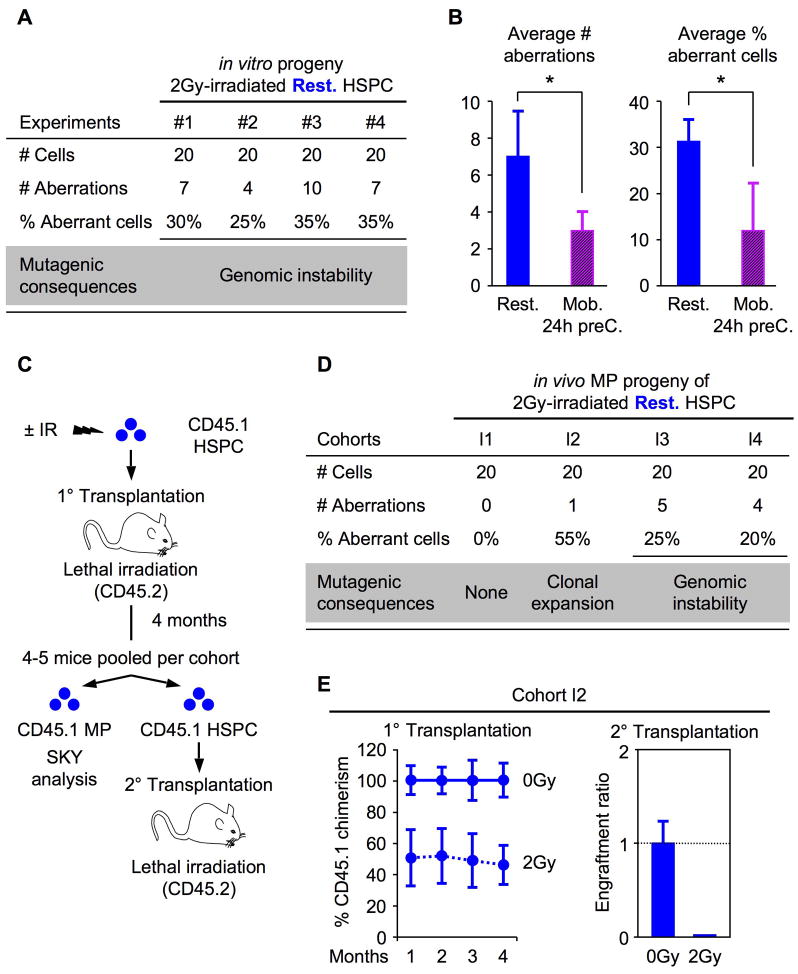
Most adult stem cells, including hematopoietic stem cells (HSCs), are maintained in a quiescent or resting state in vivo. Quiescence is widely considered to be an essential protective mechanism for stem cells that minimizes endogenous stress caused by cellular respiration and DNA replication. We demonstrate that HSC quiescence can also have detrimental effects. We found that HSCs have unique cell-intrinsic mechanisms ensuring their survival in response to ionizing irradiation (IR), which include enhanced prosurvival gene expression and strong activation of p53-mediated DNA damage response. We show that quiescent and proliferating HSCs are equally radioprotected but use different types of DNA repair mechanisms. We describe how nonhomologous end joining (NHEJ)-mediated DNA repair in quiescent HSCs is associated with acquisition of genomic rearrangements, which can persist in vivo and contribute to hematopoietic abnormalities. Our results demonstrate that quiescence is a double-edged sword that renders HSCs intrinsically vulnerable to mutagenesis following DNA damage.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Differential DNA damage response in stem and progenitor cells.Cell Stem Cell. 2010 Aug 6;7(2):145-7. doi: 10.1016/j.stem.2010.07.006. Cell Stem Cell. 2010. PMID: 20682442
-
Stem cells and DNA damage: persist or perish?Cell. 2010 Aug 6;142(3):360-2. doi: 10.1016/j.cell.2010.07.030. Cell. 2010. PMID: 20691895
References
-
- Allan JM, Travis LB. Mechanisms of therapy-related carcinogenesis. Nat Rev Cancer. 2005;5:943–955. - PubMed
-
- Bonnet D, Dick JE. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat Med. 1997;3:730–737. - PubMed
-
- Chambers SM, Goodell MA. Hematopoietic stem cell aging: wrinkles in stem cell potential. Stem Cell Rev. 2007;3:201–211. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
