

Eicosanoid pathways regulate adaptive immunity to Mycobacterium tuberculosis
- PMID: 20622882
- PMCID: PMC3150169
- DOI: 10.1038/ni.1904
Eicosanoid pathways regulate adaptive immunity to Mycobacterium tuberculosis
Abstract
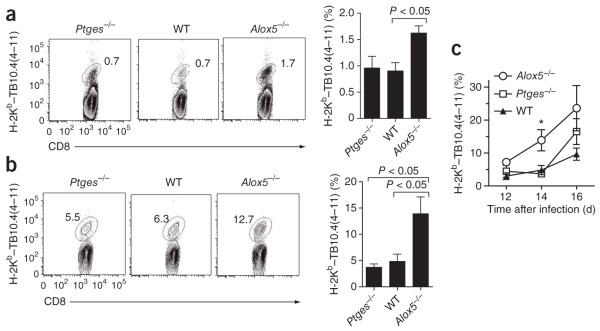
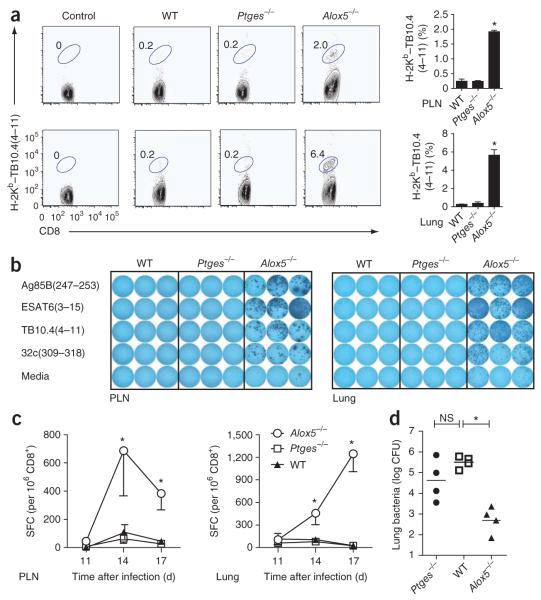
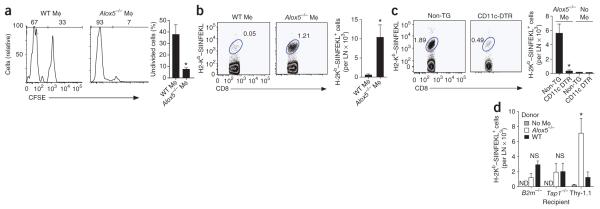
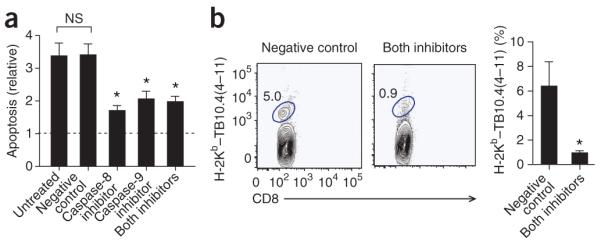
The fate of infected macrophages has an essential role in protection against Mycobacterium tuberculosis by regulating innate and adaptive immunity. M. tuberculosis exploits cell necrosis to exit from macrophages and spread. In contrast, apoptosis, which is characterized by an intact plasma membrane, is an innate mechanism that results in lower bacterial viability. Virulent M. tuberculosis inhibits apoptosis and promotes necrotic cell death by inhibiting production of prostaglandin E(2). Here we show that by activating the 5-lipoxygenase pathway, M. tuberculosis not only inhibited apoptosis but also prevented cross-presentation of its antigens by dendritic cells, which impeded the initiation of T cell immunity. Our results explain why T cell priming in response to M. tuberculosis is delayed and emphasize the importance of early immunity.
Figures

References
-
- Bhatt K, Salgame P. Host innate immune response to Mycobacterium tuberculosis. J. Clin. Immunol. 2007;27:347–362. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
