

Divergent actions of the pyrethroid insecticides S-bioallethrin, tefluthrin, and deltamethrin on rat Na(v)1.6 sodium channels
- PMID: 20624410
- PMCID: PMC2929565
- DOI: 10.1016/j.taap.2010.07.001
Divergent actions of the pyrethroid insecticides S-bioallethrin, tefluthrin, and deltamethrin on rat Na(v)1.6 sodium channels
Abstract
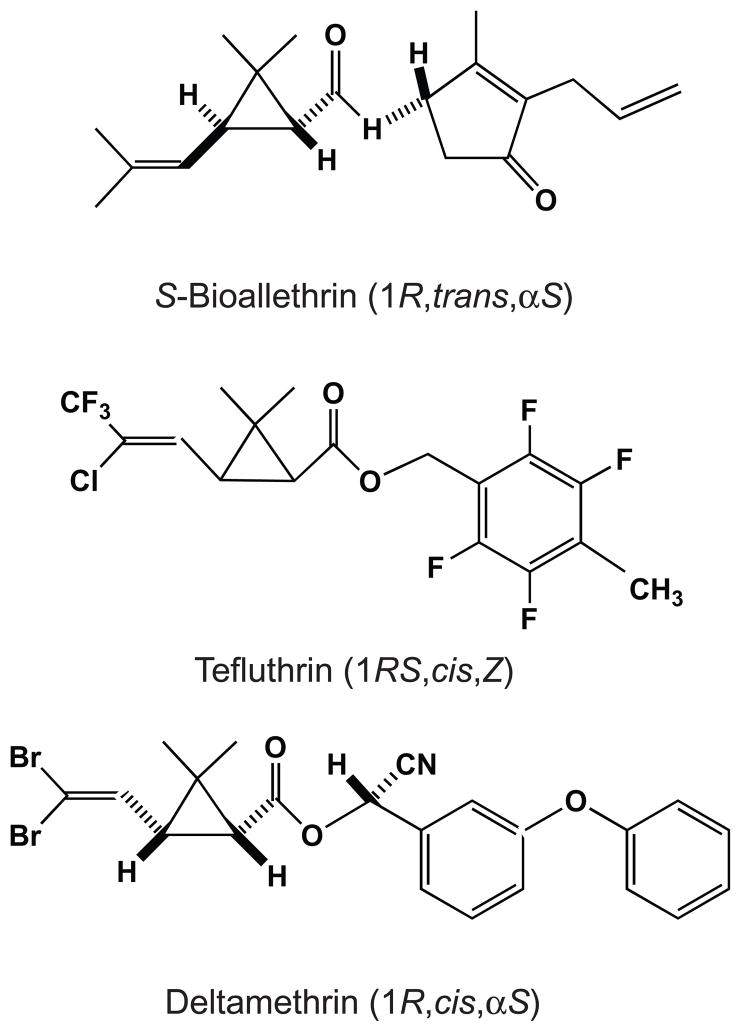
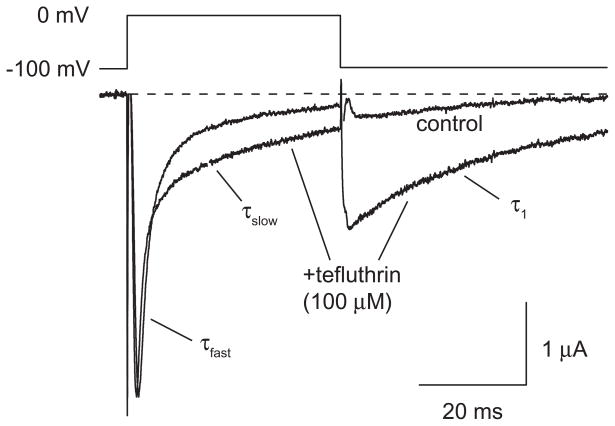
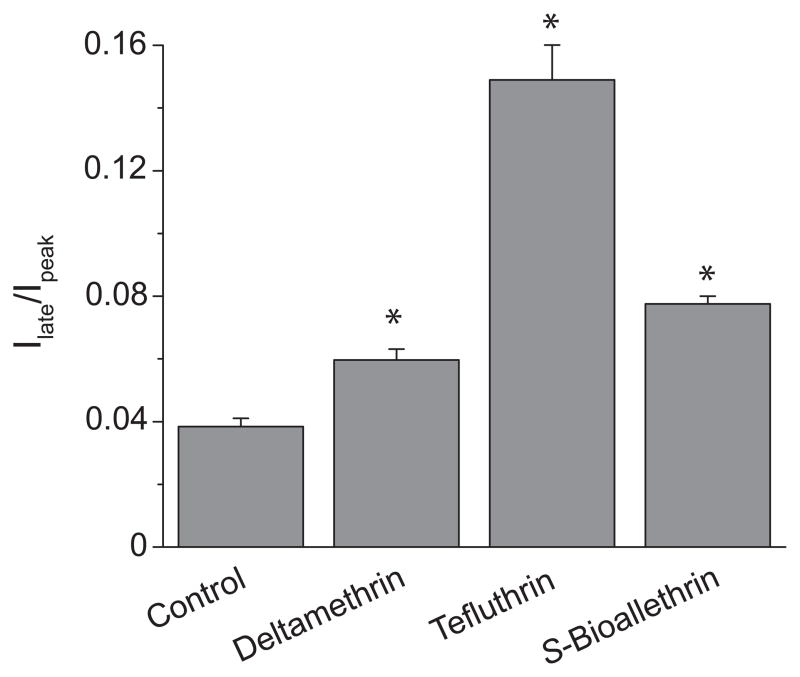
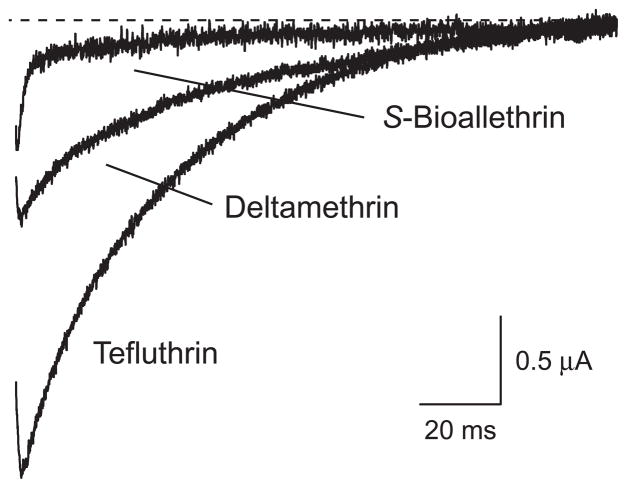
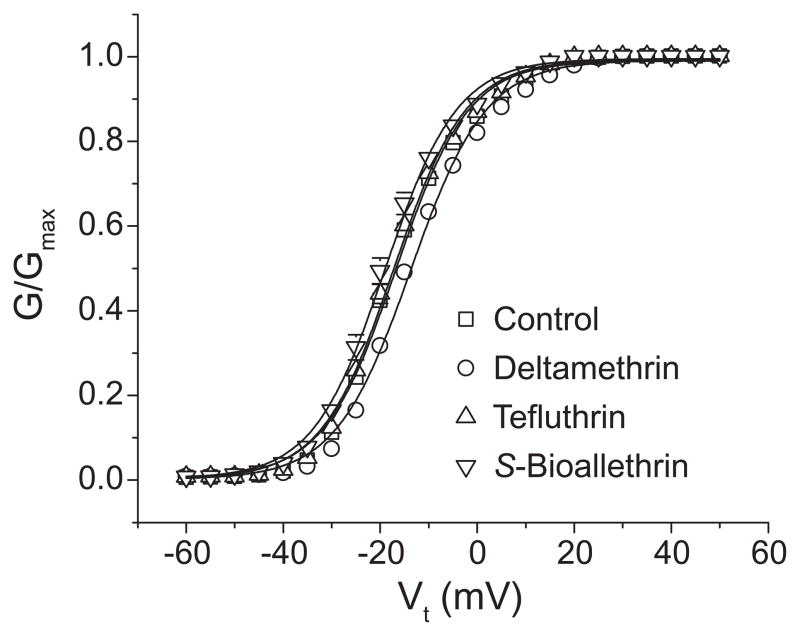
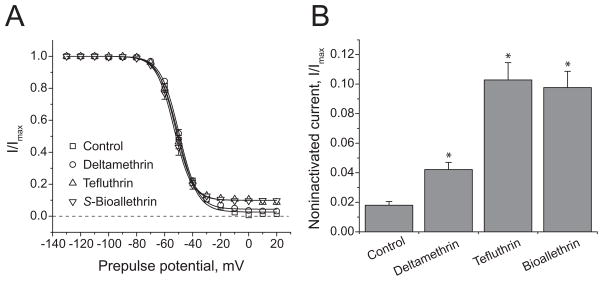
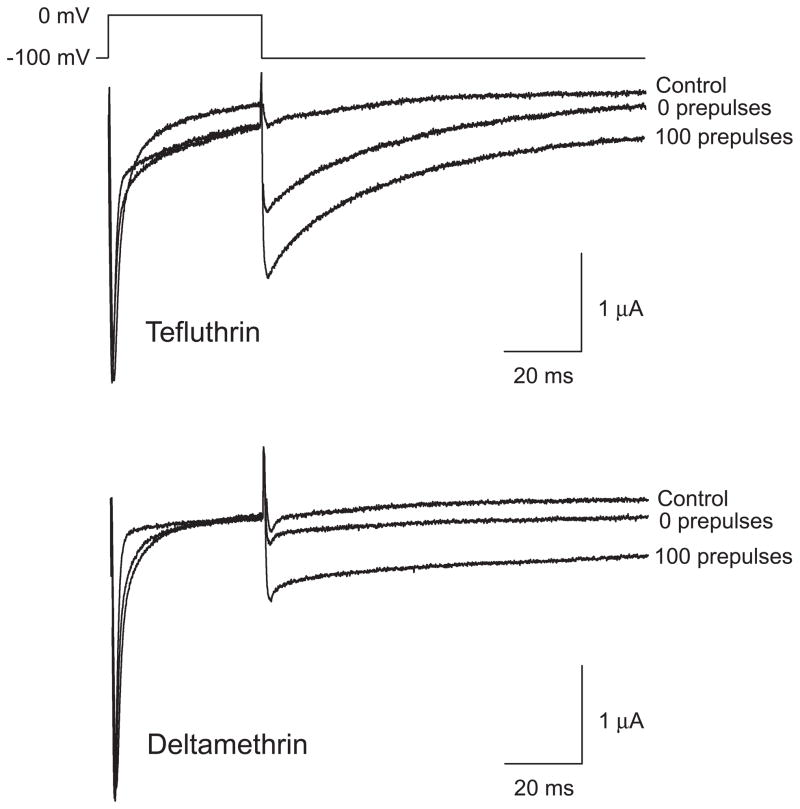
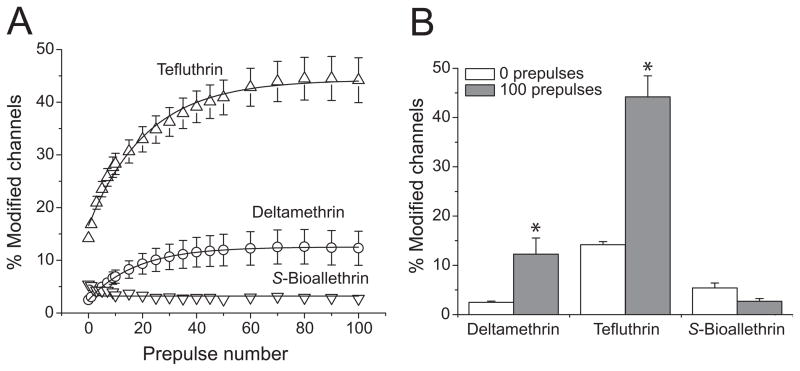
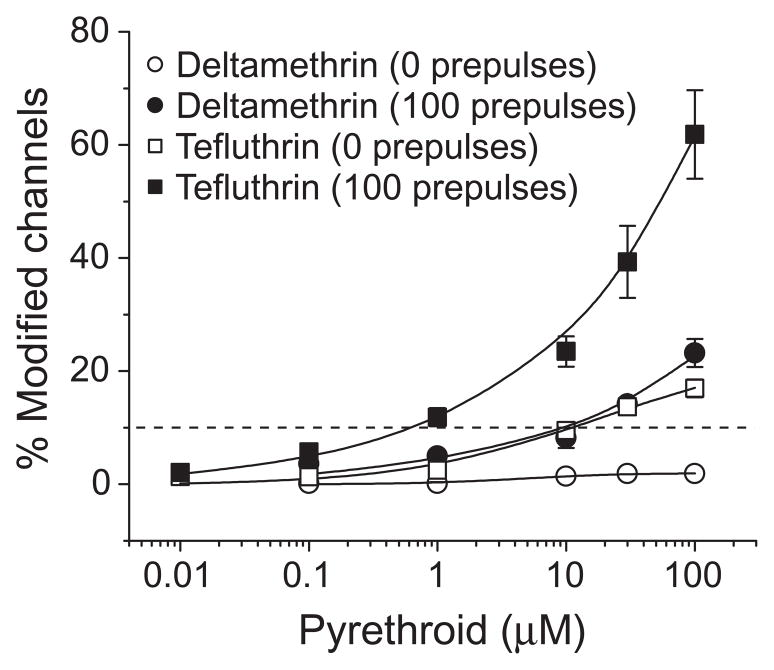
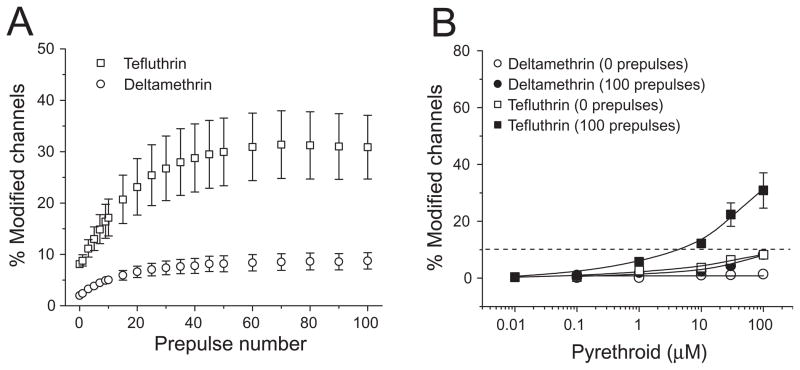
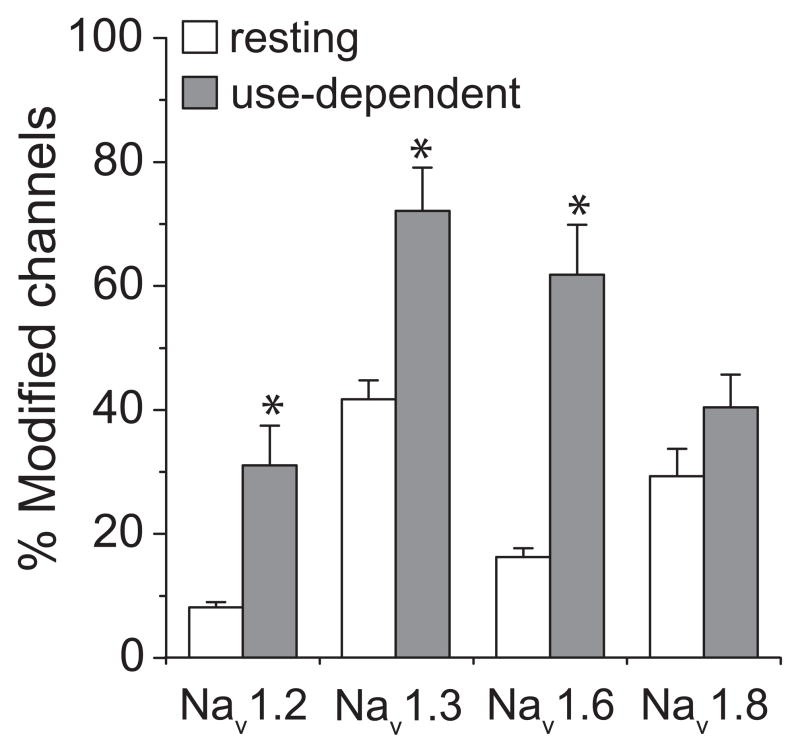
We expressed rat Na(v)1.6 sodium channels in combination with the rat beta(1) and beta(2) auxiliary subunits in Xenopus laevis oocytes and evaluated the effects of the pyrethroid insecticides S-bioallethrin, deltamethrin, and tefluthrin on expressed sodium currents using the two-electrode voltage clamp technique. S-Bioallethrin, a type I structure, produced transient modification evident in the induction of rapidly decaying sodium tail currents, weak resting modification (5.7% modification at 100 microM), and no further enhancement of modification upon repetitive activation by high-frequency trains of depolarizing pulses. By contrast deltamethrin, a type II structure, produced sodium tail currents that were ~9-fold more persistent than those caused by S-bioallethrin, barely detectable resting modification (2.5% modification at 100 microM), and 3.7-fold enhancement of modification upon repetitive activation. Tefluthrin, a type I structure with high mammalian toxicity, exhibited properties intermediate between S-bioallethrin and deltamethrin: intermediate tail current decay kinetics, much greater resting modification (14.1% at 100 microM), and 2.8-fold enhancement of resting modification upon repetitive activation. Comparison of concentration-effect data showed that repetitive depolarization increased the potency of tefluthrin approximately 15-fold and that tefluthrin was approximately 10-fold more potent than deltamethrin as a use-dependent modifier of Na(v)1.6 sodium channels. Concentration-effect data from parallel experiments with the rat Na(v)1.2 sodium channel coexpressed with the rat beta(1) and beta(2) subunits in oocytes showed that the Na(v)1.6 isoform was at least 15-fold more sensitive to tefluthrin and deltamethrin than the Na(v)1.2 isoform. These results implicate sodium channels containing the Na(v)1.6 isoform as potential targets for the central neurotoxic effects of pyrethroids.
2010 Elsevier Inc. All rights reserved.
Conflict of interest statement
Neither J. Tan nor D. M. Soderlund have conflicts of interest regarding the research described in this manuscript.
Figures
References
-
- Auld VJ, Goldin AL, Krafte DS, Marshall J, Dunn JM, Catterall WA, Lester HA, Davidson N, Dunn RJ. A rat brain Na+ channel α subunit with novel gating properties. Neuron. 1988;1:449–461. - PubMed
-
- Bloomquist JR. Neuroreceptor mechanisms in pyrethroid mode of action and resistance. In: Roe M, Kuhr RJ, editors. Reviews in Pesticide Toxicology. Toxicology Communications; Raleigh, NC: 1993a. pp. 181–226.
-
- Bloomquist JR. Toxicology, mode of action and target site-mediated resistance to insecticides acting on chloride channels. Comp Biochem Physiol. 1993b;106C:301–314. - PubMed
-
- Breckenridge CB, Holden L, Sturgess N, Weiner M, Sheets L, Sargent D, Soderlund DM, Choi JS, Symington S, Clark JM, Burr S, Ray D. Evidence for a separate mechanism of toxicity for the Type I and Type II pyrethroid insecticides. Neurotoxicology. 2009;30:S17–S31. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
