

Participation of KCNQ (Kv7) potassium channels in myogenic control of cerebral arterial diameter
- PMID: 20624791
- PMCID: PMC2976022
- DOI: 10.1113/jphysiol.2010.192823
Participation of KCNQ (Kv7) potassium channels in myogenic control of cerebral arterial diameter
Abstract
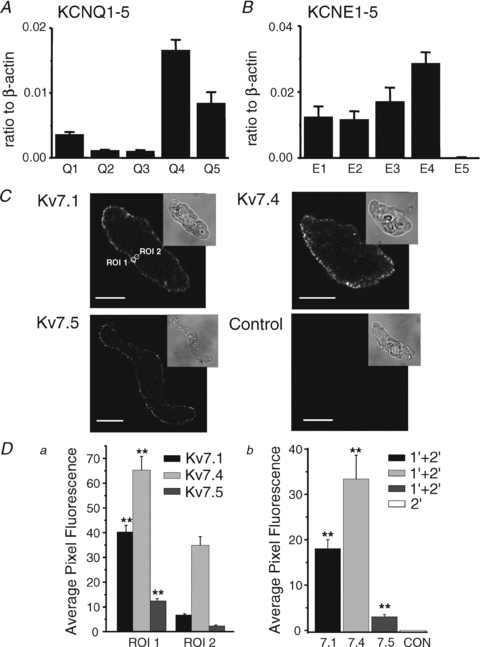
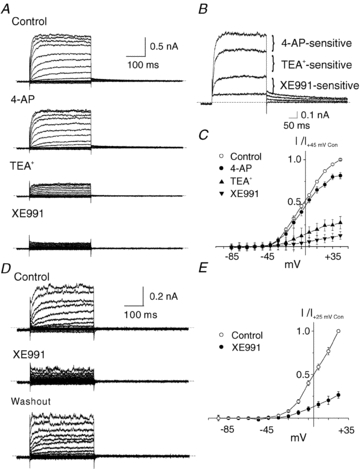
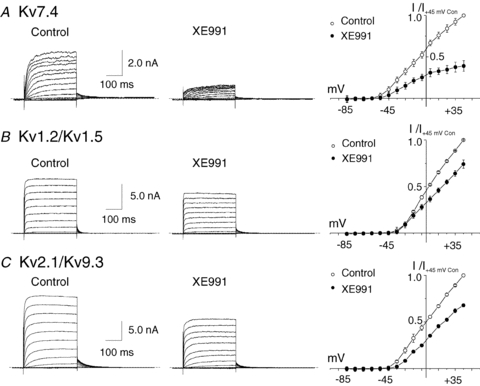
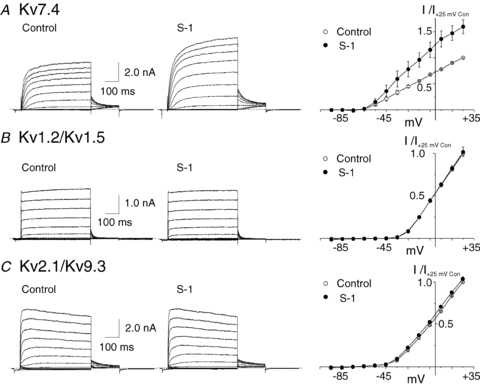
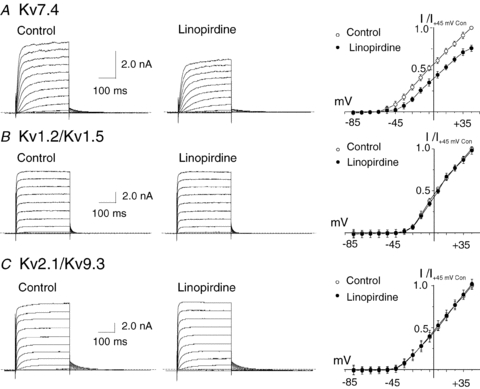
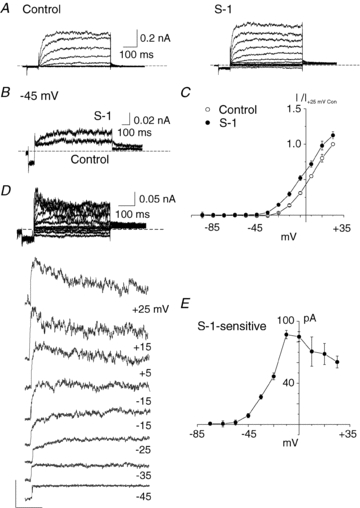
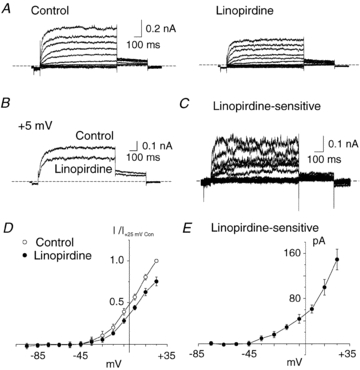
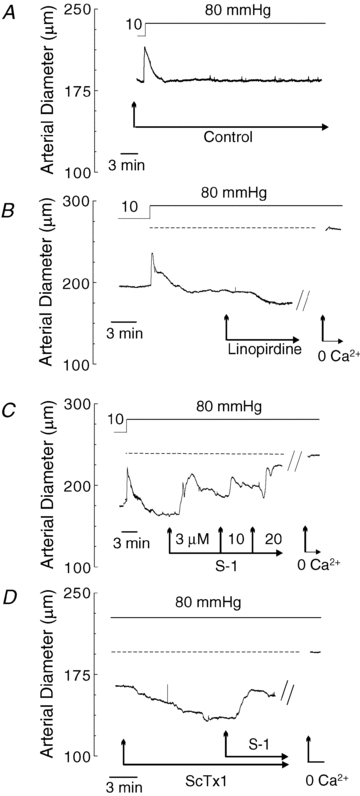
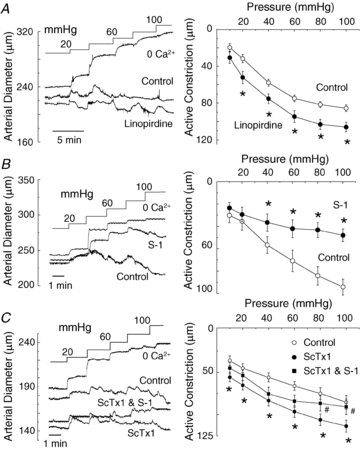
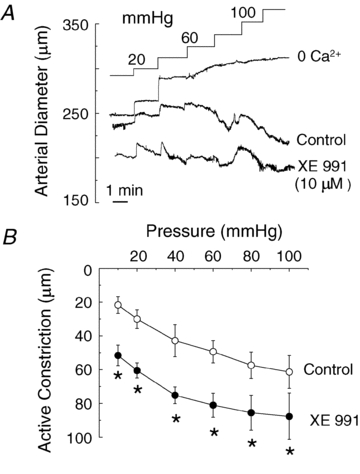
KCNQ gene expression was previously shown in various rodent blood vessels, where the products of KCNQ4 and KCNQ5, Kv7.4 and Kv7.5 potassium channel subunits, respectively, have an influence on vascular reactivity. The aim of this study was to determine if small cerebral resistance arteries of the rat express KCNQ genes and whether Kv7 channels participate in the regulation of myogenic control of diameter. Quantitative reverse transcription polymerase chain reaction (QPCR) was undertaken using RNA isolated from rat middle cerebral arteries (RMCAs) and immunocytochemistry was performed using Kv7 subunit-specific antibodies and freshly isolated RMCA myocytes. KCNQ4 message was more abundant than KCNQ5 = KCNQ1, but KCNQ2 and KCNQ3 message levels were negligible. Kv7.1, Kv7.4 and Kv7.5 immunoreactivity was present at the sarcolemma of freshly isolated RMCA myocytes. Linopirdine (1 microm) partially depressed, whereas the Kv7 activator S-1 (3 and/or 20 microm) enhanced whole-cell Kv7.4 (in HEK 293 cells), as well as native RMCA myocyte Kv current amplitude. The effects of S-1 were voltage-dependent, with progressive loss of stimulation at potentials of >15 mV. At the concentrations employed linopirdine and S-1 did not alter currents due to recombinant Kv1.2/Kv1.5 or Kv2.1/Kv9.3 channels (in HEK 293 cells) that are also expressed by RMCA myocytes. In contrast, another widely used Kv7 blocker, XE991 (10 microm), significantly attenuated native Kv current and also reduced Kv1.2/Kv1.5 and Kv2.1/Kv9.3 currents. Pressurized arterial myography was performed using RMCAs exposed to intravascular pressures of 10-100 mmHg. Linopirdine (1 microm) enhanced the myogenic response at 20 mmHg, whereas the activation of Kv7 channels with S-1 (20 microm) inhibited myogenic constriction at >20 mmHg and reversed the increased myogenic response produced by suppression of Kv2-containing channels with 30 nm stromatoxin (ScTx1). These data reveal a novel contribution of KCNQ gene products to the regulation of myogenic control of cerebral arterial diameter and suggest that Kv7 channel activating drugs may be appropriate candidates for the development of an effective therapy to ameliorate cerebral vasospasm.
Figures
References
-
- Amberg GC, Santana LF. Kv2 channels oppose myogenic constriction of rat cerebral arteries. Am J Physiol Cell Physiol. 2006;291:C348–C356. - PubMed
-
- Bentzen BH, Schmitt N, Calloe K, Dalby Brown W, Grunnet M, Olesen SP. The acrylamide (S)-1 differentially affects Kv7 (KCNQ) potassium channels. Neuropharmacology. 2006;51:1068–1077. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous