

Oxidation of Helix-3 methionines precedes the formation of PK resistant PrP
- PMID: 20625387
- PMCID: PMC2895666
- DOI: 10.1371/journal.ppat.1000977
Oxidation of Helix-3 methionines precedes the formation of PK resistant PrP
Erratum in
-
Correction: Oxidation of Helix-3 Methionines Precedes the Formation of PK Resistant PrPSc.PLoS Pathog. 2017 May 3;13(5):e1006293. doi: 10.1371/journal.ppat.1006293. eCollection 2017 May. PLoS Pathog. 2017. PMID: 28467506 Free PMC article.
Retraction in
-
Retraction: Oxidation of Helix-3 Methionines Precedes the Formation of PK Resistant PrPSc.PLoS Pathog. 2024 Dec 11;20(12):e1012775. doi: 10.1371/journal.ppat.1012775. eCollection 2024 Dec. PLoS Pathog. 2024. PMID: 39661612 Free PMC article. No abstract available.
Abstract
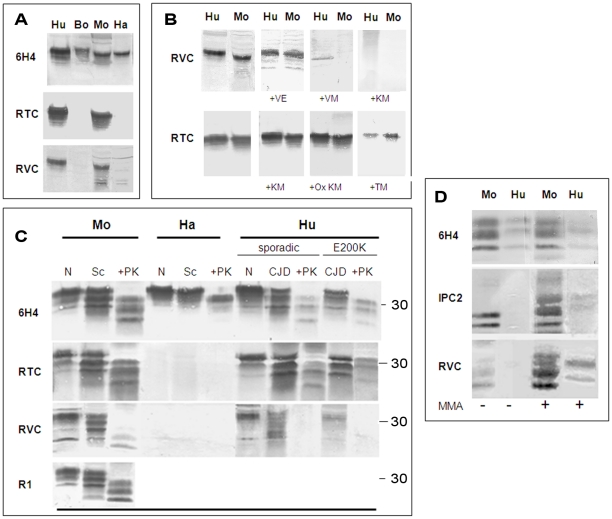
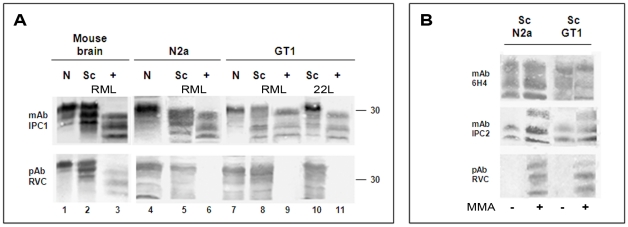
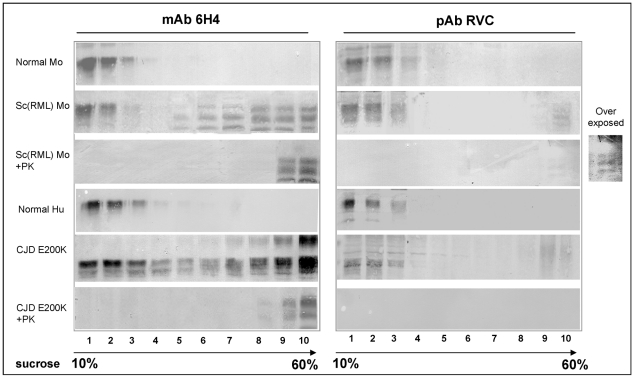
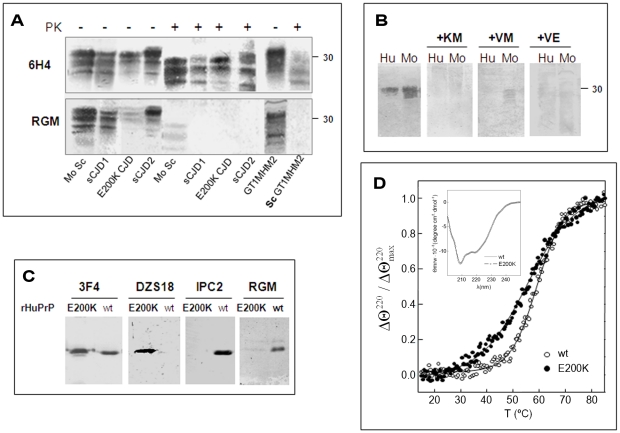
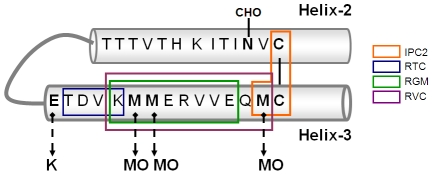
While elucidating the peculiar epitope of the alpha-PrP mAb IPC2, we found that PrPSc exhibits the sulfoxidation of residue M213 as a covalent signature. Subsequent computational analysis predicted that the presence of sulfoxide groups at both Met residues 206 and 213 destabilize the alpha-fold, suggesting oxidation may facilitate the conversion of PrPC into PrPSc. To further study the effect of oxidation on prion formation, we generated pAbs to linear PrP peptides encompassing the Helix-3 region, as opposed to the non-linear complexed epitope of IPC2. We now show that pAbs, whose epitopes comprise Met residues, readily detected PrPC, but could not recognize most PrPSc bands unless they were vigorously reduced. Next, we showed that the alpha-Met pAbs did not recognize newly formed PrPSc, as is the case for the PK resistant PrP present in lines of prion infected cells. In addition, these reagents did not detect intermediate forms such as PK sensitive and partially aggregated PrPs present in infected brains. Finally, we show that PrP molecules harboring the pathogenic mutation E200K, which is linked to the most common form of familial CJD, may be spontaneously oxidized. We conclude that the oxidation of methionine residues in Helix-3 represents an early and important event in the conversion of PrPC to PrPSc. We believe that further investigation into the mechanism and role of PrP oxidation will be central in finally elucidating the mechanism by which a normal cell protein converts into a pathogenic entity that causes fatal brain degeneration.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Prusiner SB, Scott MR, DeArmond SJ, Cohen FE. Prion protein biology. Cell. 1998;93(3):337–348. - PubMed
-
- Brown DR. Neurodegeneration and oxidative stress: prion disease results from loss of antioxidant defence. Folia Neuropathol. 2005;43:229–243. - PubMed
-
- Nadal RC, Abdelraheim SR, Brazier MW, Rigby SE, Brown DR, et al. Prion protein does not redox-silence Cu2+, but is a sacrificial quencher of hydroxyl radicals. Free Radic Biol Med. 2007;42:79–89. - PubMed
-
- Brown DR, Clive C, Haswell SJ. Antioxidant activity related to copper binding of native prion protein. J Neurochem. 2001;76:69–76. - PubMed
-
- Brown DR, Sassoon J. Copper-dependent functions for the prion protein. Mol Biotechnol. 2002;22:165–178. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
