

A model of Costeff Syndrome reveals metabolic and protective functions of mitochondrial OPA3
- PMID: 20627962
- PMCID: PMC2927703
- DOI: 10.1242/dev.043745
A model of Costeff Syndrome reveals metabolic and protective functions of mitochondrial OPA3
Abstract
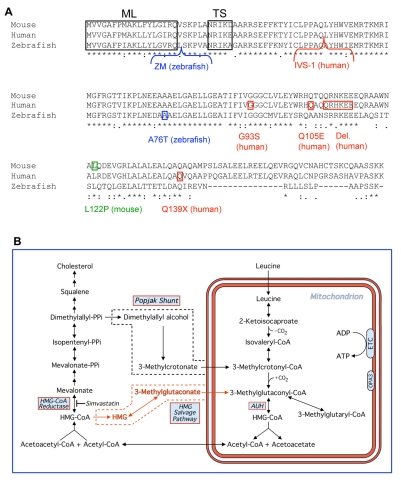
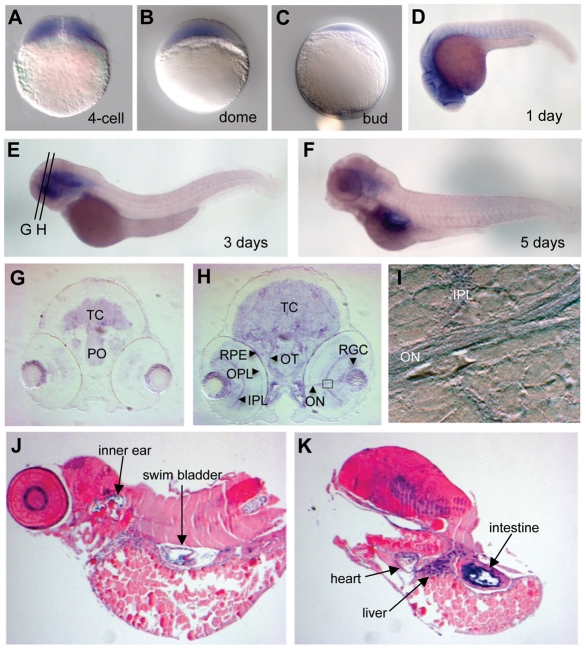
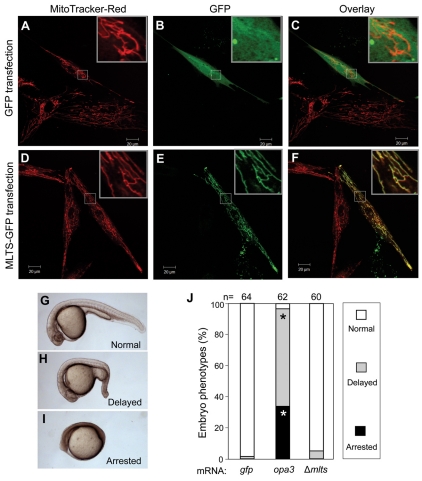
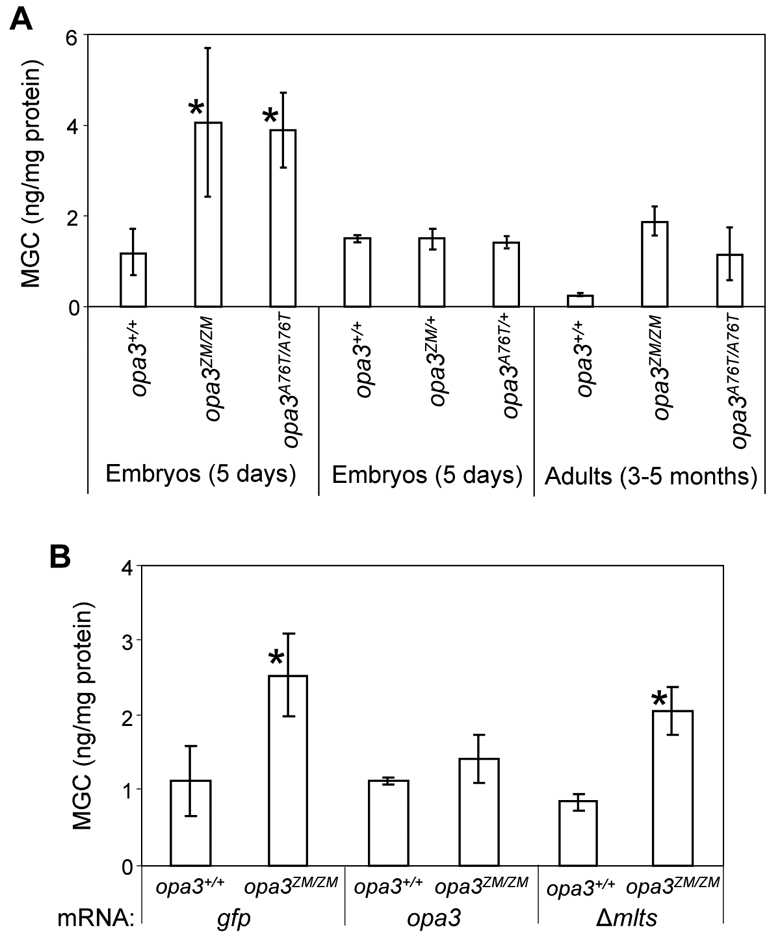
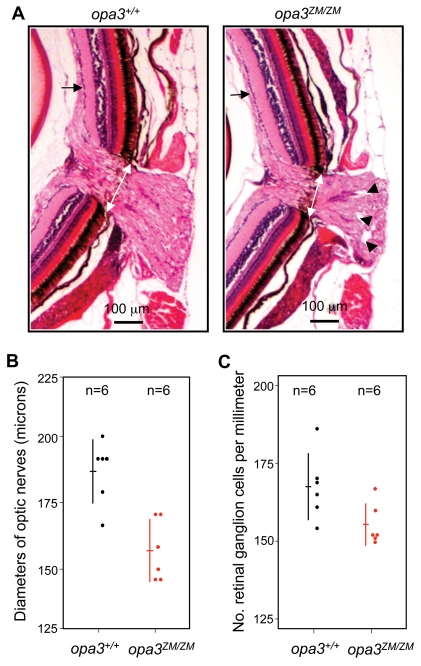
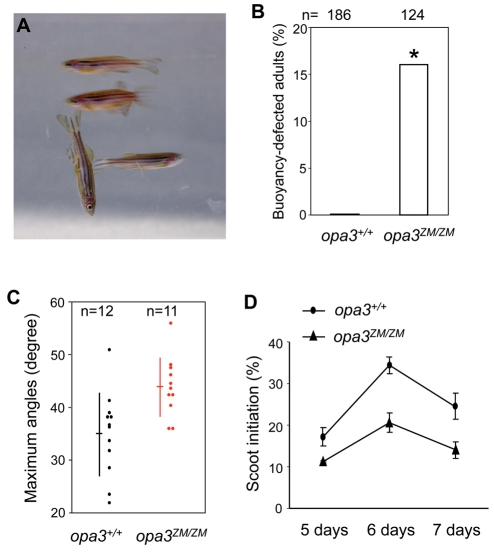
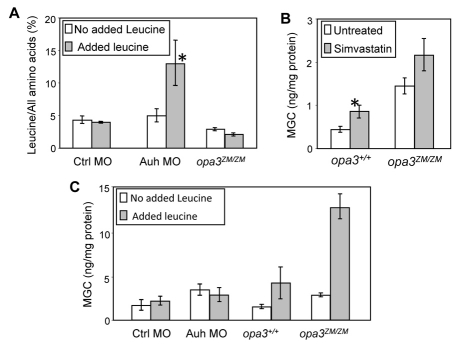
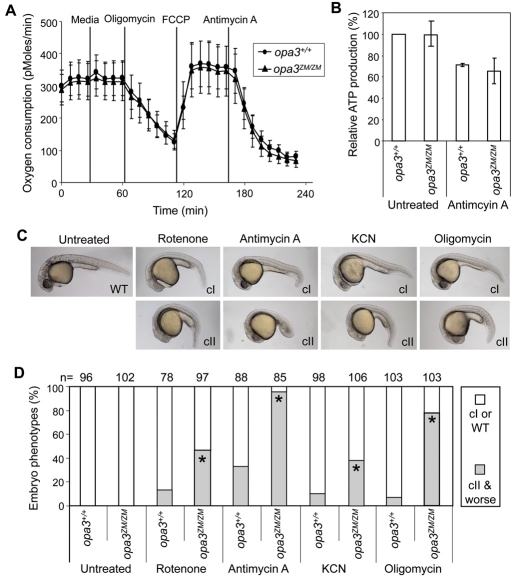
Costeff Syndrome, which is caused by mutations in the OPTIC ATROPHY 3 (OPA3) gene, is an early-onset syndrome characterized by urinary excretion of 3-methylglutaconic acid (MGC), optic atrophy and movement disorders, including ataxia and extrapyramidal dysfunction. The OPA3 protein is enriched in the inner mitochondrial membrane and has mitochondrial targeting signals, but a requirement for mitochondrial localization has not been demonstrated. We find zebrafish opa3 mRNA to be expressed in the optic nerve and retinal layers, the counterparts of which in humans have high mitochondrial activity. Transcripts of zebrafish opa3 are also expressed in the embryonic brain, inner ear, heart, liver, intestine and swim bladder. We isolated a zebrafish opa3 null allele for which homozygous mutants display increased MGC levels, optic nerve deficits, ataxia and an extrapyramidal movement disorder. This correspondence of metabolic, ophthalmologic and movement abnormalities between humans and zebrafish demonstrates a phylogenetic conservation of OPA3 function. We also find that delivery of exogenous Opa3 can reduce increased MGC levels in opa3 mutants, and this reduction requires the mitochondrial localization signals of Opa3. By manipulating MGC precursor availability, we infer that elevated MGC in opa3 mutants derives from extra-mitochondrial HMG-CoA through a non-canonical pathway. The opa3 mutants have normal mitochondrial oxidative phosphorylation profiles, but are nonetheless sensitive to inhibitors of the electron transport chain, which supports clinical recommendations that individuals with Costeff Syndrome avoid mitochondria-damaging agents. In summary, this paper introduces a faithful Costeff Syndrome model and demonstrates a requirement for mitochondrial OPA3 to limit HMG-CoA-derived MGC and protect the electron transport chain against inhibitory compounds.
Figures
References
-
- Bernhardt R. R., Tongiorgi E., Anzini P., Schachner M. (1996). Increased expression of specific recognition molecules by retinal ganglion cells and by optic pathway glia accompanies the successful regeneration of retinal axons in adult zebrafish. J. Comp. Neurol. 376, 253-253 - PubMed
-
- Bione S., D'Adamo P., Maestrini E., Gedeon A. K., Bolhuis P. A., Toniolo D. (1996). A novel X-linked gene, G4.5. is responsible for Barth syndrome. Nat. Genet. 12, 385-385 - PubMed
-
- Burgess H. A., Granato M. (2007). Modulation of locomotor activity in larval zebrafish during light adaptation. J. Exp. Biol. 210, 2526-2526 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
