

Estrogen modification of human glutamate dehydrogenases is linked to enzyme activation state
- PMID: 20628048
- PMCID: PMC2951212
- DOI: 10.1074/jbc.M110.146084
Estrogen modification of human glutamate dehydrogenases is linked to enzyme activation state
Abstract
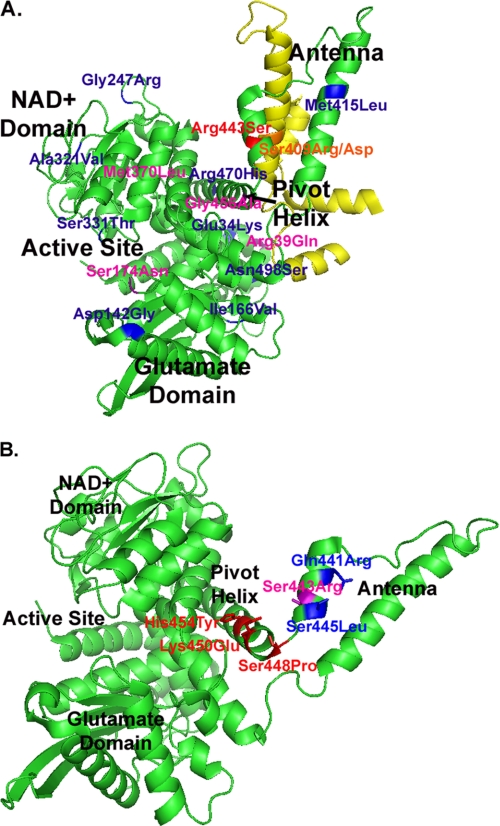
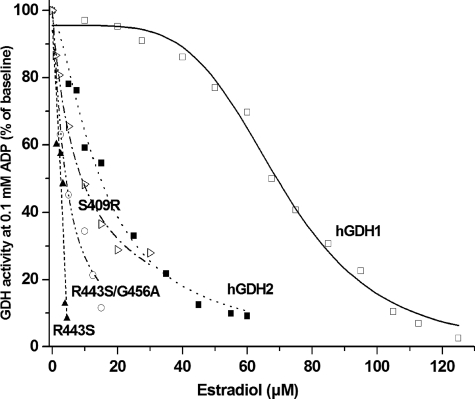
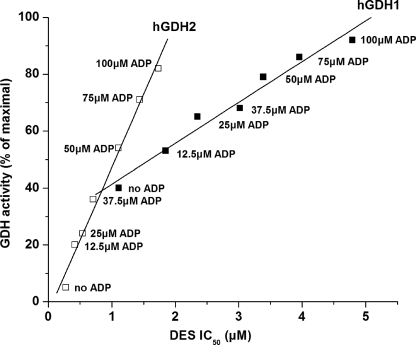
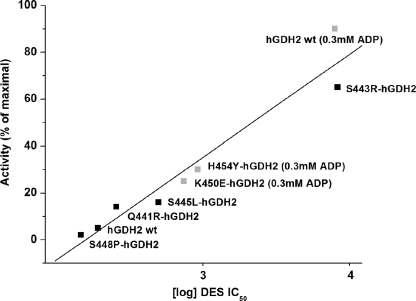
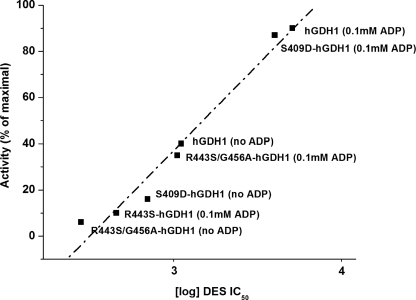
Mammalian glutamate dehydrogenase (GDH) is a housekeeping enzyme central to the metabolism of glutamate. Its activity is potently inhibited by GTP (IC(50) = 0.1-0.3 μM) and thought to be controlled by the need of the cell in ATP. Estrogens are also known to inhibit mammalian GDH, but at relatively high concentrations. Because, in addition to this housekeeping human (h) GDH1, humans have acquired via a duplication event an hGDH2 isoform expressed in human cortical astrocytes, we tested here the interaction of estrogens with the two human isoenzymes. The results showed that, under base-line conditions, diethylstilbestrol potently inhibited hGDH2 (IC(50) = 0.08 ± 0.01 μM) and with ∼18-fold lower affinity hGDH1 (IC(50) = 1.67 ± 0.06 μM; p < 0.001). Similarly, 17β-estradiol showed a ∼18-fold higher affinity for hGDH2 (IC(50) = 1.53 ± 0.24 μM) than for hGDH1 (IC(50) = 26.94 ± 1.07 μM; p < 0.001). Also, estriol and progesterone were more potent inhibitors of hGDH2 than hGDH1. Structure/function analyses revealed that the evolutionary R443S substitution, which confers low basal activity, was largely responsible for sensitivity of hGDH2 to estrogens. Inhibition of both human GDHs by estrogens was inversely related to their state of activation induced by ADP, with the slope of this correlation being steeper for hGDH2 than for hGDH1. Also, the study of hGDH1 and hGDH2 mutants displaying different states of activation revealed that the affinity of estrogen for these enzymes correlated inversely (R = 0.99; p = 0.0001) with basal catalytic activity. Because astrocytes are known to synthesize estrogens, these hormones, by interacting potently with hGDH2 in its closed state, may contribute to regulation of glutamate metabolism in brain.
Figures
References
-
- Gruber C. J., Tschugguel W., Schneeberger C., Huber J. C. (2002) N. Engl. J. Med. 346, 340–352 - PubMed
-
- Green P. S., Simpkins J. W. (2000) Int. J. Dev. Neurosci. 18, 347–358 - PubMed
-
- Amantea D., Russo R., Bagetta G., Corasaniti M. T. (2005) Pharmacol. Res. 52, 119–132 - PubMed
-
- Ba F., Pang P. K., Benishin C. G. (2004) Neurochem. Int. 45, 31–38 - PubMed
-
- Goodman Y., Bruce A. J., Cheng B., Mattson M. P. (1996) J. Neurochem. 66, 1836–1844 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
