

TGFbeta/BMP type I receptors ALK1 and ALK2 are essential for BMP9-induced osteogenic signaling in mesenchymal stem cells
- PMID: 20628059
- PMCID: PMC2937990
- DOI: 10.1074/jbc.M110.130518
TGFbeta/BMP type I receptors ALK1 and ALK2 are essential for BMP9-induced osteogenic signaling in mesenchymal stem cells
Abstract
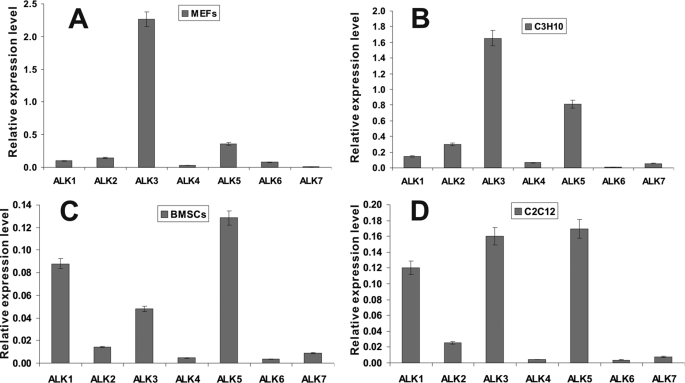
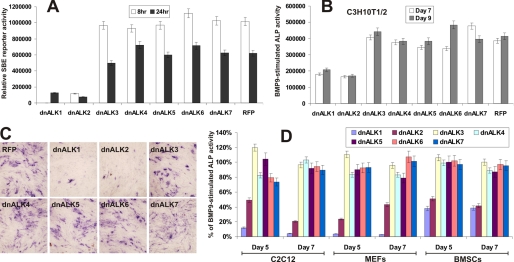
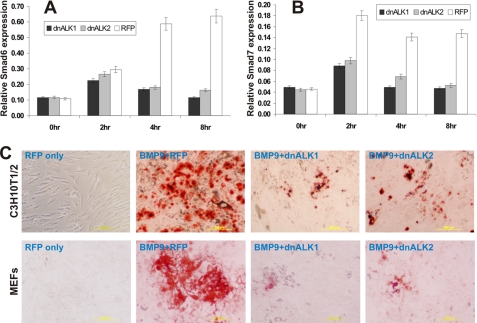
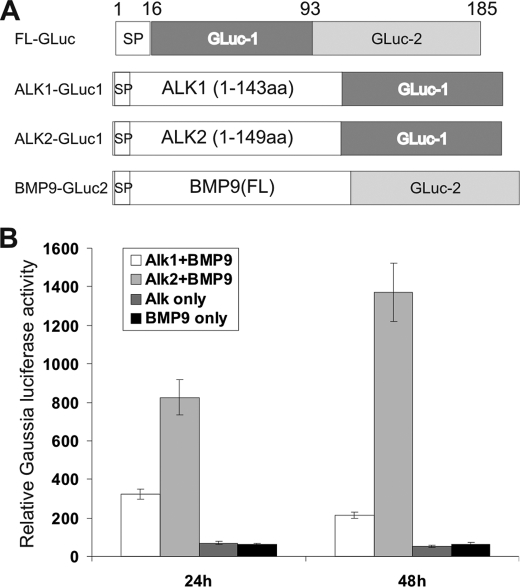
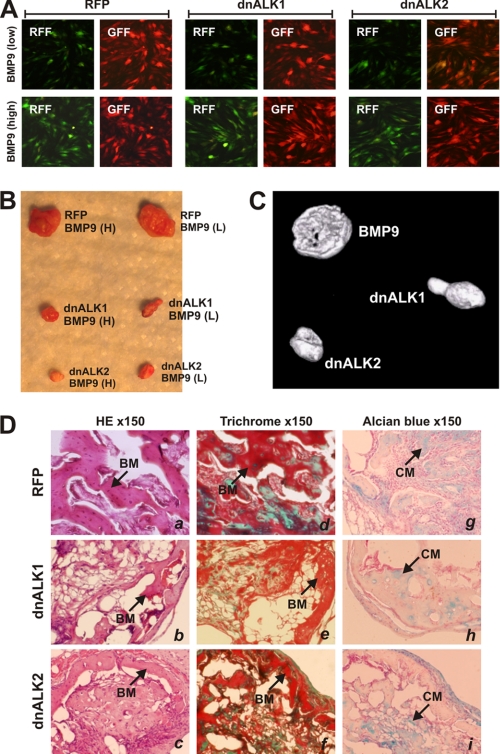
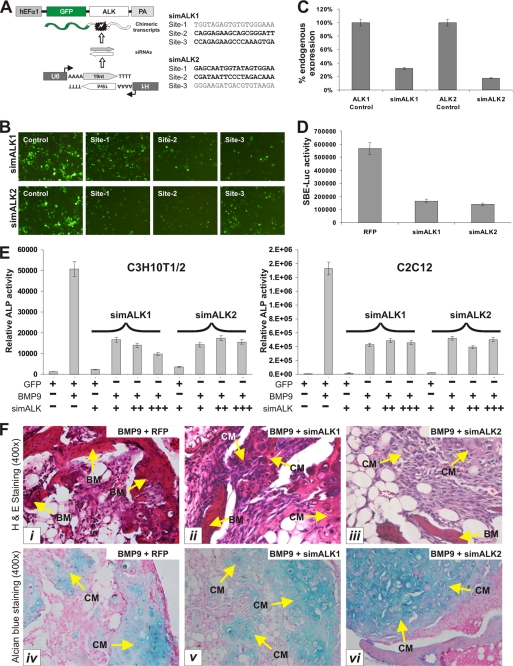
Mesenchymal stem cells (MSCs) are bone marrow stromal cells that can differentiate into multiple lineages. We previously demonstrated that BMP9 is one of the most potent BMPs to induce osteogenic differentiation of MSCs. BMP9 is one of the least studied BMPs. Whereas ALK1, ALK5, and/or endoglin have recently been reported as potential BMP9 type I receptors in endothelial cells, little is known about type I receptor involvement in BMP9-induced osteogenic differentiation in MSCs. Here, we conduct a comprehensive analysis of the functional role of seven type I receptors in BMP9-induced osteogenic signaling in MSCs. We have found that most of the seven type I receptors are expressed in MSCs. However, using dominant-negative mutants for the seven type I receptors, we demonstrate that only ALK1 and ALK2 mutants effectively inhibit BMP9-induced osteogenic differentiation in vitro and ectopic ossification in MSC implantation assays. Protein fragment complementation assays demonstrate that ALK1 and ALK2 directly interact with BMP9. Likewise, RNAi silencing of ALK1 and ALK2 expression inhibits BMP9-induced BMPR-Smad activity and osteogenic differentiation in MSCs both in vitro and in vivo. Therefore, our results strongly suggest that ALK1 and ALK2 may play an important role in mediating BMP9-induced osteogenic differentiation. These findings should further aid us in understanding the molecular mechanism through which BMP9 regulates osteogenic differentiation of MSCs.
Figures


Similar articles
-
Fibroblast growth factor 2 inhibits bone morphogenetic protein 9-induced osteogenic differentiation of mesenchymal stem cells by repressing Smads signaling and subsequently reducing Smads dependent up-regulation of ALK1 and ALK2.Int J Biochem Cell Biol. 2013 Aug;45(8):1639-46. doi: 10.1016/j.biocel.2013.05.005. Epub 2013 May 13. Int J Biochem Cell Biol. 2013. PMID: 23680673
-
Schnurri-3 regulates BMP9-induced osteogenic differentiation and angiogenesis of human amniotic mesenchymal stem cells through Runx2 and VEGF.Cell Death Dis. 2020 Jan 29;11(1):72. doi: 10.1038/s41419-020-2279-5. Cell Death Dis. 2020. Retraction in: Cell Death Dis. 2021 Dec 9;12(12):1144. doi: 10.1038/s41419-021-04443-8. PMID: 31996667 Free PMC article. Retracted.
-
Retinoic acids potentiate BMP9-induced osteogenic differentiation of mesenchymal progenitor cells.PLoS One. 2010 Jul 30;5(7):e11917. doi: 10.1371/journal.pone.0011917. PLoS One. 2010. PMID: 20689834 Free PMC article.
-
Bone morphogenetic proteins.Growth Factors. 2004 Dec;22(4):233-41. doi: 10.1080/08977190412331279890. Growth Factors. 2004. PMID: 15621726 Review.
-
Regulation of the ALK1 ligands, BMP9 and BMP10.Biochem Soc Trans. 2016 Aug 15;44(4):1135-41. doi: 10.1042/BST20160083. Biochem Soc Trans. 2016. PMID: 27528761 Review.
Cited by
-
Cross-talk between EGF and BMP9 signalling pathways regulates the osteogenic differentiation of mesenchymal stem cells.J Cell Mol Med. 2013 Sep;17(9):1160-72. doi: 10.1111/jcmm.12097. Epub 2013 Jul 11. J Cell Mol Med. 2013. PMID: 23844832 Free PMC article.
-
P38 and ERK1/2 MAPKs act in opposition to regulate BMP9-induced osteogenic differentiation of mesenchymal progenitor cells.PLoS One. 2012;7(8):e43383. doi: 10.1371/journal.pone.0043383. Epub 2012 Aug 17. PLoS One. 2012. Retraction in: PLoS One. 2018 Nov 2;13(11):e0207157. doi: 10.1371/journal.pone.0207157. PMID: 22912865 Free PMC article. Retracted.
-
Terbium promotes adhesion and osteogenic differentiation of mesenchymal stem cells via activation of the Smad-dependent TGF-β/BMP signaling pathway.J Biol Inorg Chem. 2014 Aug;19(6):879-91. doi: 10.1007/s00775-014-1119-4. Epub 2014 Mar 2. J Biol Inorg Chem. 2014. PMID: 24585101
-
The Prodomain-Containing BMP9 Produced from a Stable Line Effectively Regulates the Differentiation of Mesenchymal Stem Cells.Int J Med Sci. 2016 Jan 1;13(1):8-18. doi: 10.7150/ijms.13333. eCollection 2016. Int J Med Sci. 2016. PMID: 26816490 Free PMC article.
-
Osteogenic differentiation cues of the bone morphogenetic protein-9 (BMP-9) and its recent advances in bone tissue regeneration.Mater Sci Eng C Mater Biol Appl. 2021 Jan;120:111748. doi: 10.1016/j.msec.2020.111748. Epub 2020 Nov 27. Mater Sci Eng C Mater Biol Appl. 2021. PMID: 33545890 Free PMC article. Review.
References
-
- Prockop D. J. (1997) Science 276, 71–74 - PubMed
-
- Pittenger M. F., Mackay A. M., Beck S. C., Jaiswal R. K., Douglas R., Mosca J. D., Moorman M. A., Simonetti D. W., Craig S., Marshak D. R. (1999) Science 284, 143–147 - PubMed
-
- Luu H. H., Song W. X., Luo X., Manning D., Luo J., Deng Z. L., Sharff K. A., Montag A. G., Haydon R. C., He T. C. (2007) J. Orthop. Res. 25, 665–677 - PubMed
-
- Deng Z. L., Sharff K. A., Tang N., Song W. X., Luo J., Luo X., Chen J., Bennett E., Reid R., Manning D., Xue A., Montag A. G., Luu H. H., Haydon R. C., He T. C. (2008) Front Biosci. 13, 2001–2021 - PubMed
-
- Varga A. C., Wrana J. L. (2005) Oncogene 24, 5713–5721 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
