

Hepatic HIF-2 regulates erythropoietic responses to hypoxia in renal anemia
- PMID: 20628150
- PMCID: PMC2974609
- DOI: 10.1182/blood-2010-02-270322
Hepatic HIF-2 regulates erythropoietic responses to hypoxia in renal anemia
Abstract
The kidney is the main physiologic source of erythropoietin (EPO) in the adult and responds to decreases in tissue oxygenation with increased EPO production. Although studies in mice with liver-specific or global gene inactivation have shown that hypoxia-inducible factor 2 (Hif-2) plays a major role in the regulation of Epo during infancy and in the adult, respectively, the contribution of renal HIF-2 signaling to systemic EPO homeostasis and the role of extrarenal HIF-2 in erythropoiesis, in the absence of kidney EPO, have not been examined directly. Here, we used Cre-loxP recombination to ablate Hif-2α in the kidney, whereas Hif-2-mediated hypoxia responses in the liver and other Epo-producing tissues remained intact. We found that the hypoxic induction of renal Epo is completely Hif-2 dependent and that, in the absence of renal Hif-2, hepatic Hif-2 takes over as the main regulator of serum Epo levels. Furthermore, we provide evidence that hepatocyte-derived Hif-2 is involved in the regulation of iron metabolism genes, supporting a role for HIF-2 in the coordination of EPO synthesis with iron homeostasis.
Figures
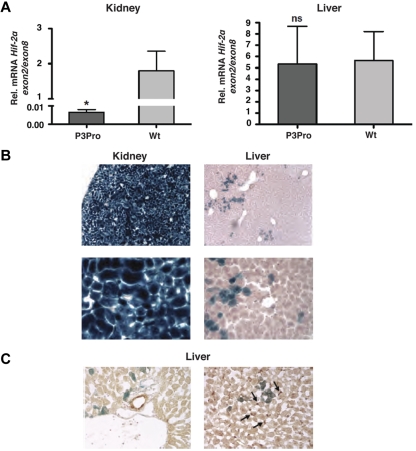
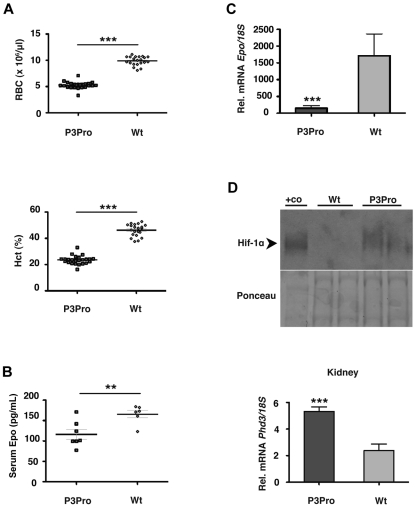
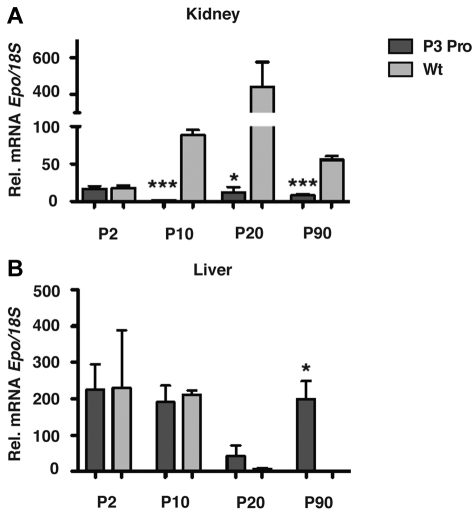
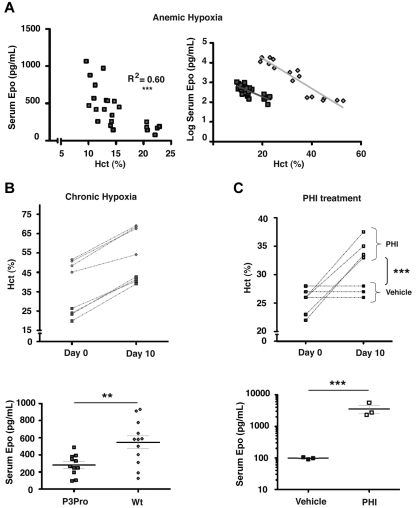
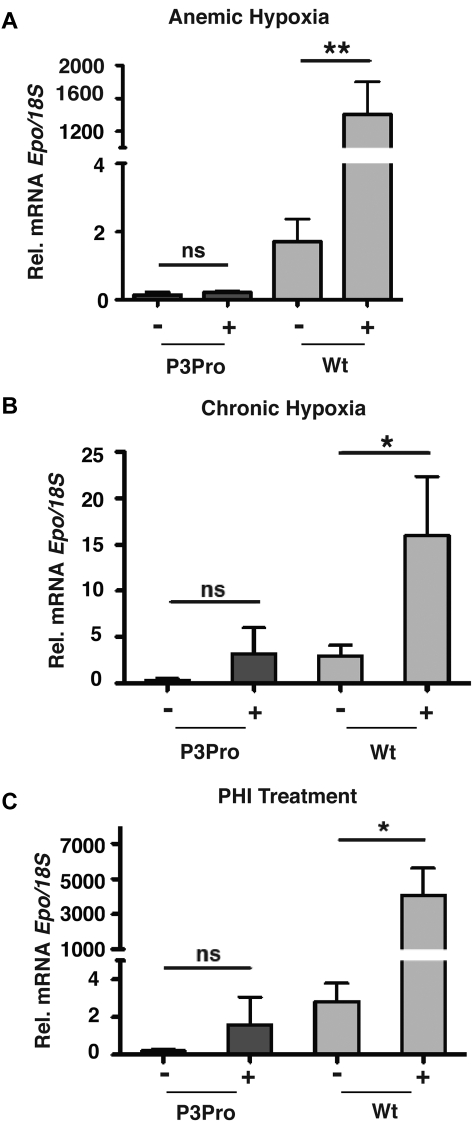
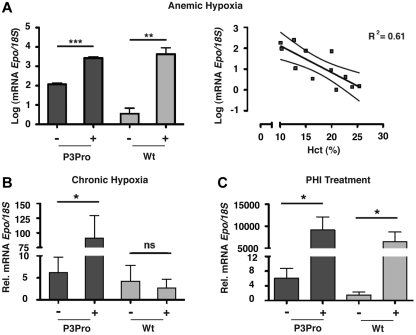
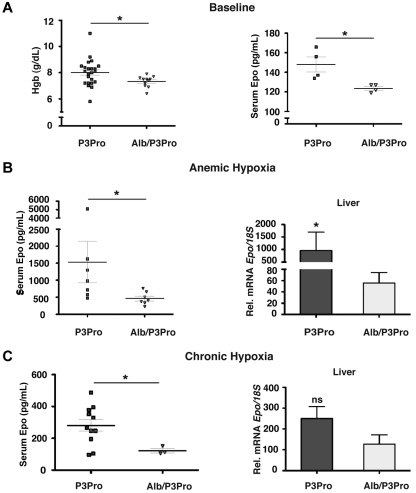
Comment in
-
Dual control: the HIF-2 regulator.Blood. 2010 Oct 21;116(16):2870-1. doi: 10.1182/blood-2010-08-297606. Blood. 2010. PMID: 20966176 No abstract available.
References
-
- Fandrey J. Oxygen-dependent and tissue-specific regulation of erythropoietin gene expression. Am J Physiol Regul Integr Comp Physiol. 2004;286(6):R977–R988. - PubMed
-
- Jelkmann W. Control of erythropoietin gene expression and its use in medicine. Methods Enzymol. 2007;435:179–197. - PubMed
-
- Wu H, Lee SH, Gao J, Liu X, Iruela-Arispe ML. Inactivation of erythropoietin leads to defects in cardiac morphogenesis. Development. 1999;126(16):3597–3605. - PubMed
-
- Koury ST, Bondurant MC, Koury MJ. Localization of erythropoietin synthesizing cells in murine kidneys by in situ hybridization. Blood. 1988;71(2):524–527. - PubMed
-
- Bachmann S, Le Hir M, Eckardt KU. Co-localization of erythropoietin mRNA and ecto-5′-nucleotidase immunoreactivity in peritubular cells of rat renal cortex indicates that fibroblasts produce erythropoietin. J Histochem Cytochem. 1993;41(3):335–341. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
