

Low doses of killed parasite in CpG elicit vigorous CD4+ T cell responses against blood-stage malaria in mice
- PMID: 20628205
- PMCID: PMC2912178
- DOI: 10.1172/JCI39222
Low doses of killed parasite in CpG elicit vigorous CD4+ T cell responses against blood-stage malaria in mice
Abstract
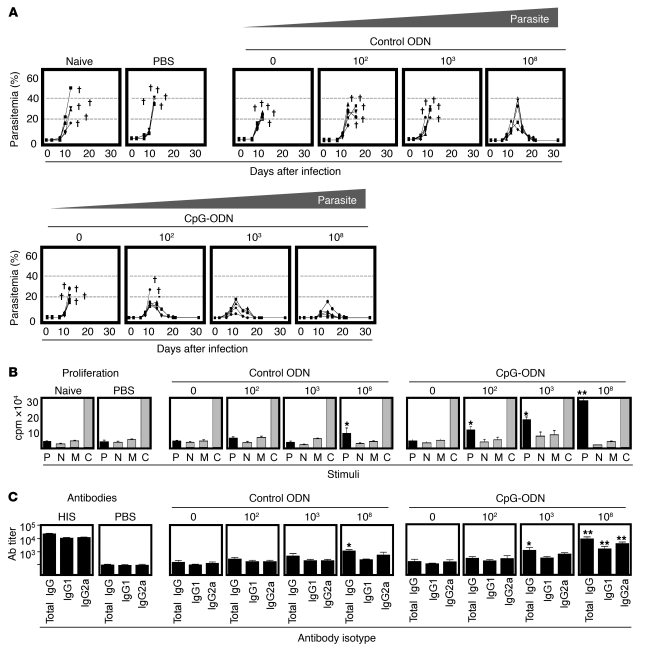
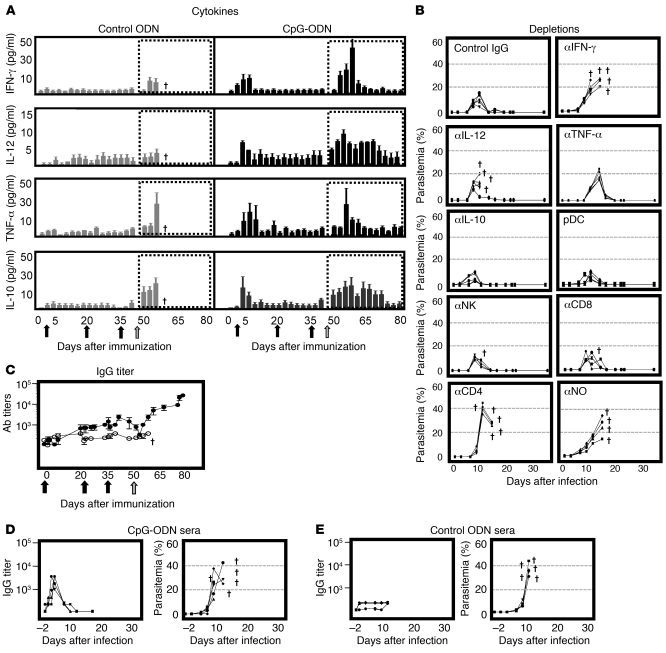
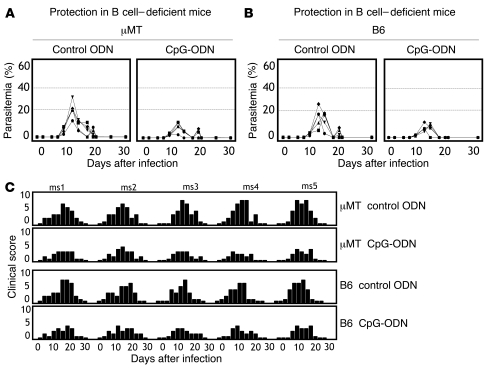
Development of a vaccine that targets blood-stage malaria parasites is imperative if we are to sustainably reduce the morbidity and mortality caused by this infection. Such a vaccine should elicit long-lasting immune responses against conserved determinants in the parasite population. Most blood-stage vaccines, however, induce protective antibodies against surface antigens, which tend to be polymorphic. Cell-mediated responses, on the other hand, offer the theoretical advantage of targeting internal antigens that are more likely to be conserved. Nonetheless, few of the current blood-stage vaccine candidates are able to harness vigorous T cell immunity. Here, we present what we believe to be a novel blood-stage whole-organism vaccine that, by combining low doses of killed parasite with CpG-oligodeoxynucleotide (CpG-ODN) adjuvant, was able to elicit strong and cross-reactive T cell responses in mice. Our data demonstrate that immunization of mice with 1,000 killed parasites in CpG-ODN engendered durable and cross-strain protection by inducing a vigorous response that was dependent on CD4+ T cells, IFN-gamma, and nitric oxide. If applicable to humans, this approach should facilitate the generation of robust, cross-reactive T cell responses against malaria as well as antigen availability for vaccine manufacture.
Figures




References
-
- Fox JP. Modes of action of poliovirus vaccines and relation to resulting immunity. Rev Infect Dis. 1984;6 suppl 2:S352–S355. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
