

Divergent cortical generators of MEG and EEG during human sleep spindles suggested by distributed source modeling
- PMID: 20628643
- PMCID: PMC2898804
- DOI: 10.1371/journal.pone.0011454
Divergent cortical generators of MEG and EEG during human sleep spindles suggested by distributed source modeling
Abstract
Background: Sleep spindles are approximately 1-second bursts of 10-15 Hz activity, occurring during normal stage 2 sleep. In animals, sleep spindles can be synchronous across multiple cortical and thalamic locations, suggesting a distributed stable phase-locked generating system. The high synchrony of spindles across scalp EEG sites suggests that this may also be true in humans. However, prior MEG studies suggest multiple and varying generators.
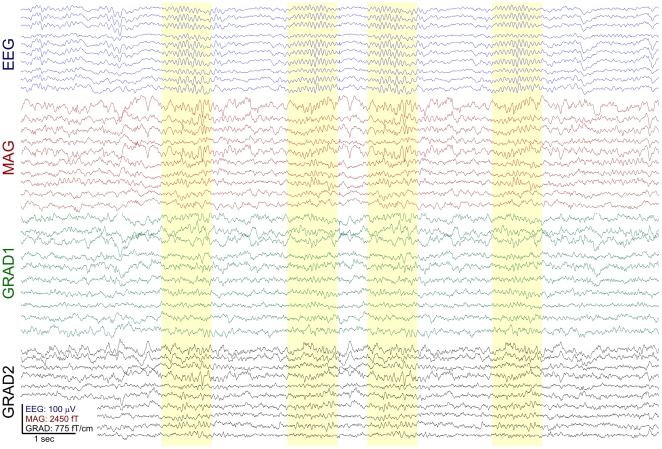
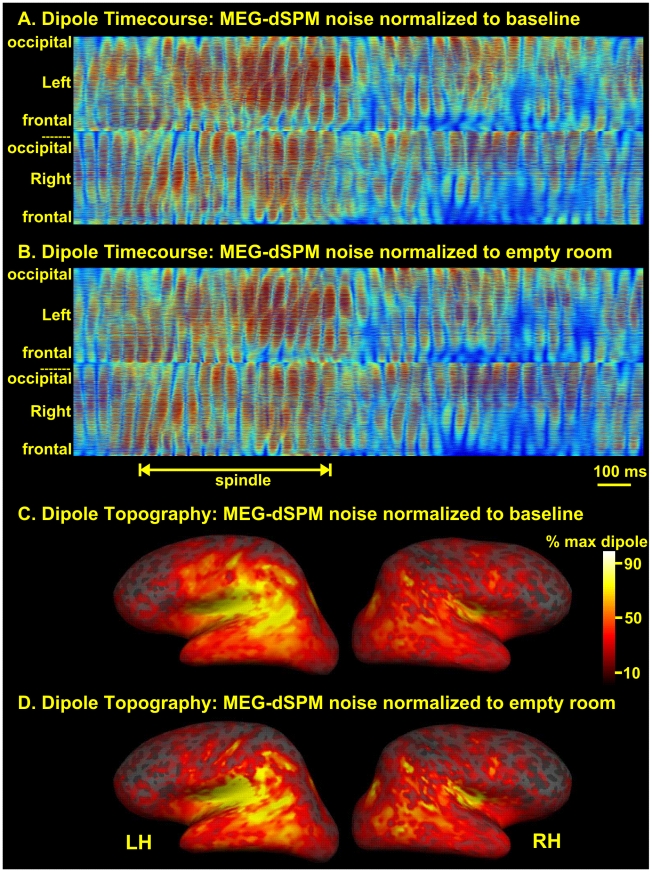
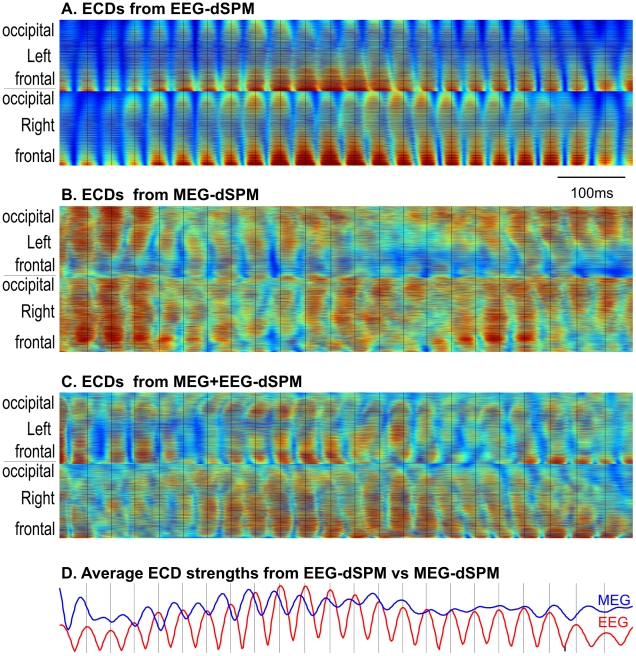
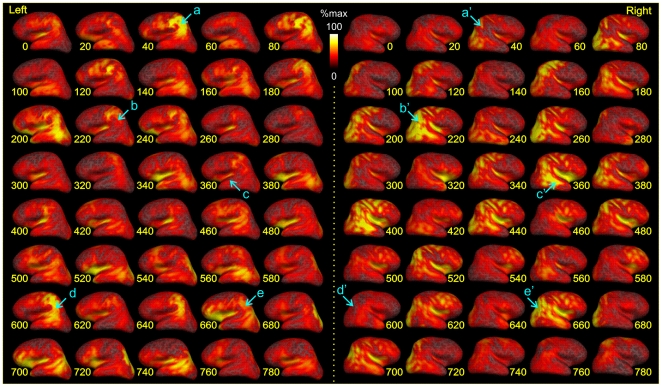
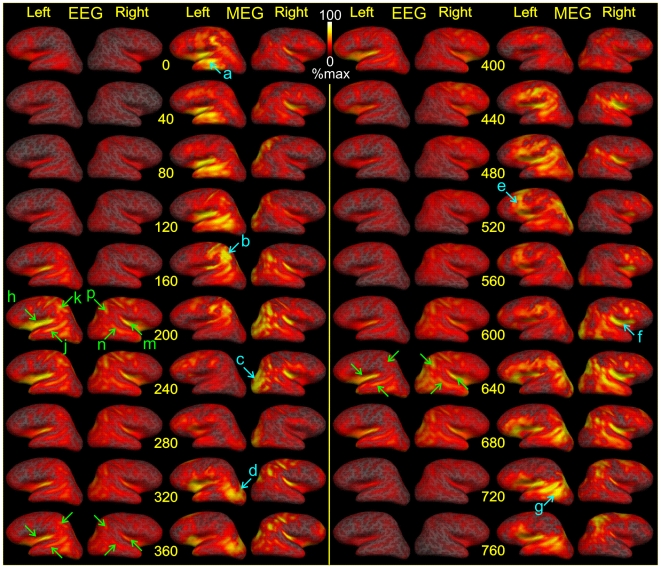
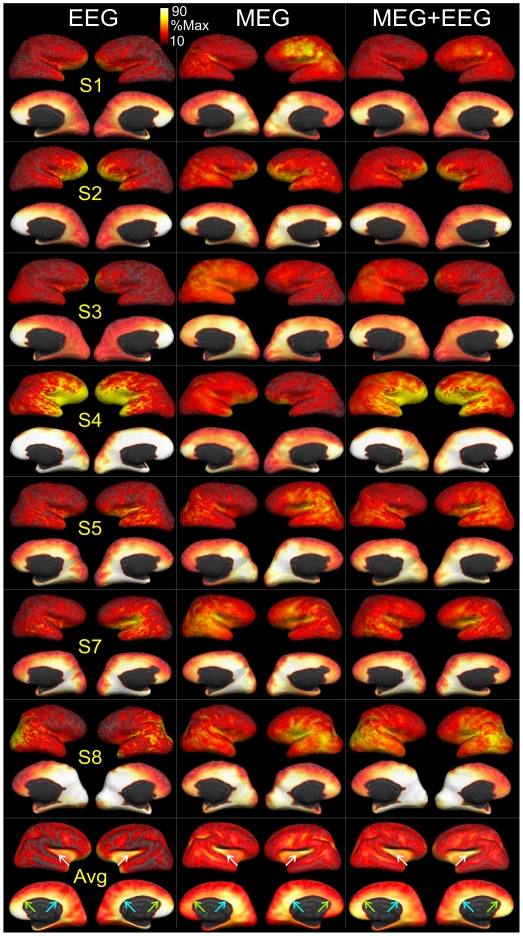
Methodology/principal findings: We recorded 306 channels of MEG simultaneously with 60 channels of EEG during naturally occurring spindles of stage 2 sleep in 7 healthy subjects. High-resolution structural MRI was obtained in each subject, to define the shells for a boundary element forward solution and to reconstruct the cortex providing the solution space for a noise-normalized minimum norm source estimation procedure. Integrated across the entire duration of all spindles, sources estimated from EEG and MEG are similar, diffuse and widespread, including all lobes from both hemispheres. However, the locations, phase and amplitude of sources simultaneously estimated from MEG versus EEG are highly distinct during the same spindles. Specifically, the sources estimated from EEG are highly synchronous across the cortex, whereas those from MEG rapidly shift in phase, hemisphere, and the location within the hemisphere.
Conclusions/significance: The heterogeneity of MEG sources implies that multiple generators are active during human sleep spindles. If the source modeling is correct, then EEG spindles are generated by a different, diffusely synchronous system. Animal studies have identified two thalamo-cortical systems, core and matrix, that produce focal or diffuse activation and thus could underlie MEG and EEG spindles, respectively. Alternatively, EEG spindles could reflect overlap at the sensors of the same sources as are seen from the MEG. Although our results generally match human intracranial recordings, additional improvements are possible and simultaneous intra- and extra-cranial measures are needed to test their accuracy.
Conflict of interest statement
Figures
References
-
- Loomis AL, Harvey EN, Hobart G. Potential rhythms of cerebral cortex during sleep. Science. 1935;81:597–598. - PubMed
-
- Gibbs FA, Gibbs EL. Cambridge: Addison-Wesley Press; 1950. Atlas of Electroencephalography.
-
- Zepelin H, Siegel JM, Tobler I. Mammalian sleep. In: Kryger MH, Roth T, Dement WC, editors. Principles and Practice of Sleep Medicine. London: Saunders; 1994. pp. 91–100.
-
- Andersen P, Andersson SA. New York: Meredith Corp; 1968. Physiological basis of the alpha rhythm.
-
- Spencer WA, Brookhart JM. A study of spontaneous spindle waves in sensorimotor cortex of cat. J Neurophysiol. 1961;24:50–65.
