

Posttranslational modification of the NH2-terminal region of CXCL5 by proteases or peptidylarginine Deiminases (PAD) differently affects its biological activity
- PMID: 20630876
- PMCID: PMC2943270
- DOI: 10.1074/jbc.M110.119388
Posttranslational modification of the NH2-terminal region of CXCL5 by proteases or peptidylarginine Deiminases (PAD) differently affects its biological activity
Abstract
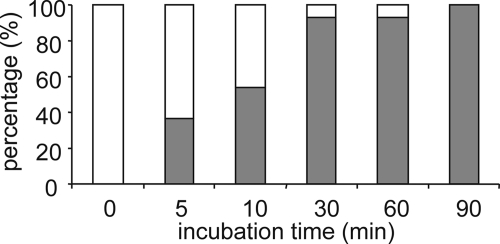
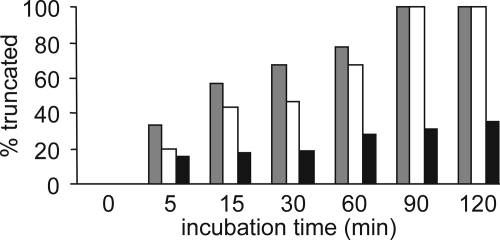
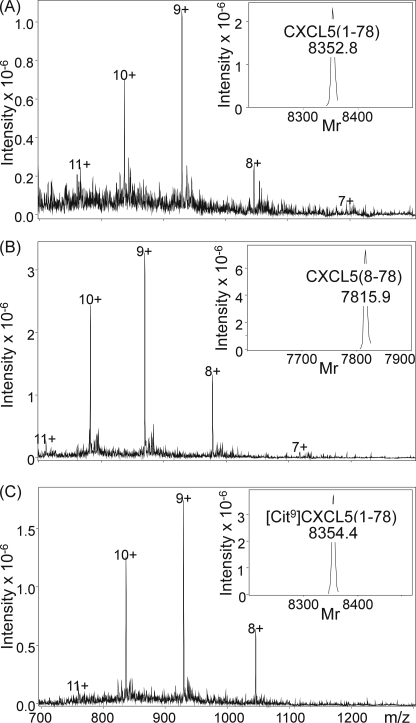
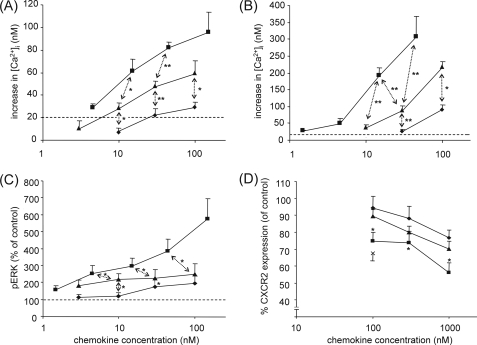
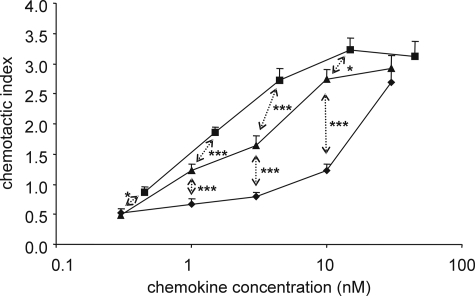
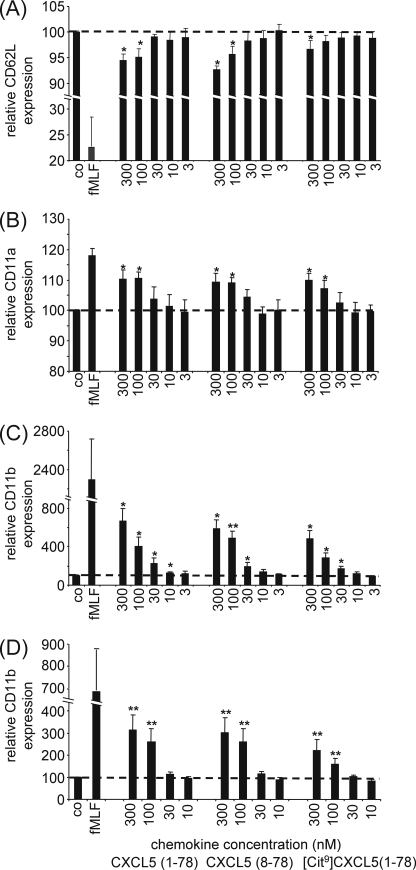
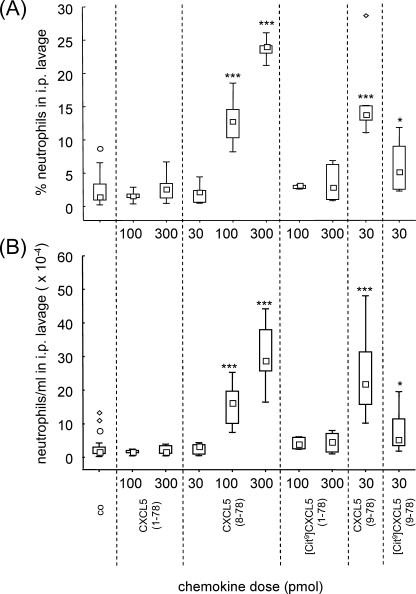
Posttranslational modifications, e.g. proteolysis, glycosylation, and citrullination regulate chemokine function, affecting leukocyte migration during inflammatory responses. Here, modification of CXCL5/epithelial cell-derived neutrophil-activating protein-78 (ENA-78) by proteases or peptidylarginine deiminases (PAD) was evaluated. Slow CXCL5(1-78) processing by the myeloid cell marker aminopeptidase N/CD13 into CXCL5(2-78) hardly affected its in vitro activity, but slowed down the activation of CXCL5 by the neutrophil protease cathepsin G. PAD, an enzyme with a potentially important function in autoimmune diseases, site-specifically deiminated Arg(9) in CXCL5 to citrulline, generating [Cit(9)]CXCL5(1-78). Compared with CXCL5(1-78), [Cit(9)]CXCL5(1-78) less efficiently induced intracellular calcium signaling, phosphorylation of extracellular signal-regulated kinase, internalization of CXCR2, and in vitro neutrophil chemotaxis. In contrast, conversion of CXCL5 into the previously reported natural isoform CXCL5(8-78) provided at least 3-fold enhanced biological activity in these tests. Citrullination, but not NH(2)-terminal truncation, reduced the capacity of CXCL5 to up-regulate the expression of the integrin α-chain CD11b on neutrophils. Truncation nor citrullination significantly affected the ability of CXCL5 to up-regulate CD11a expression or shedding of CD62L. In line with the in vitro results, CXCL5(8-78) and CXCL5(9-78) induced a more pronounced neutrophil influx in vivo compared with CXCL5(1-78). Administration of 300 pmol of either CXCL5(1-78) or [Cit(9)]CXCL5(1-78) failed to attract neutrophils to the peritoneal cavity. Citrullination of the more potent CXCL5(9-78) lowers its chemotactic potency in vivo and confirms the tempering effect of citrullination in vitro. The highly divergent effects of modifications of CXCL5 on neutrophil influx underline the potential importance of tissue-specific interactions between chemokines and PAD or proteases.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
