

Macroscopic stiffening of embryonic tissues via microtubules, RhoGEF and the assembly of contractile bundles of actomyosin
- PMID: 20630946
- PMCID: PMC2910388
- DOI: 10.1242/dev.045997
Macroscopic stiffening of embryonic tissues via microtubules, RhoGEF and the assembly of contractile bundles of actomyosin
Abstract
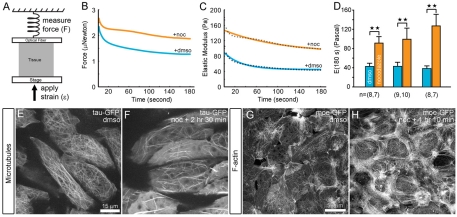
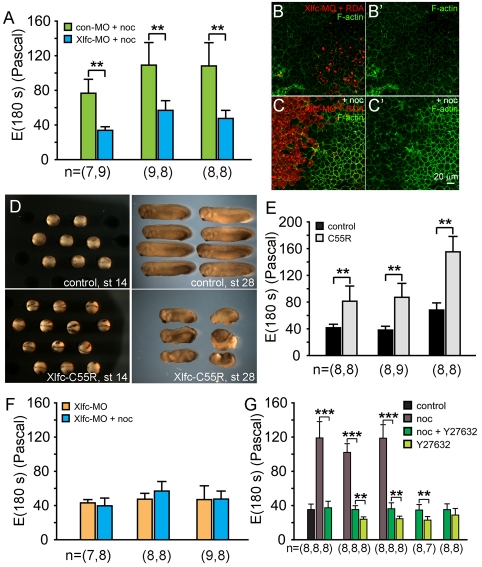
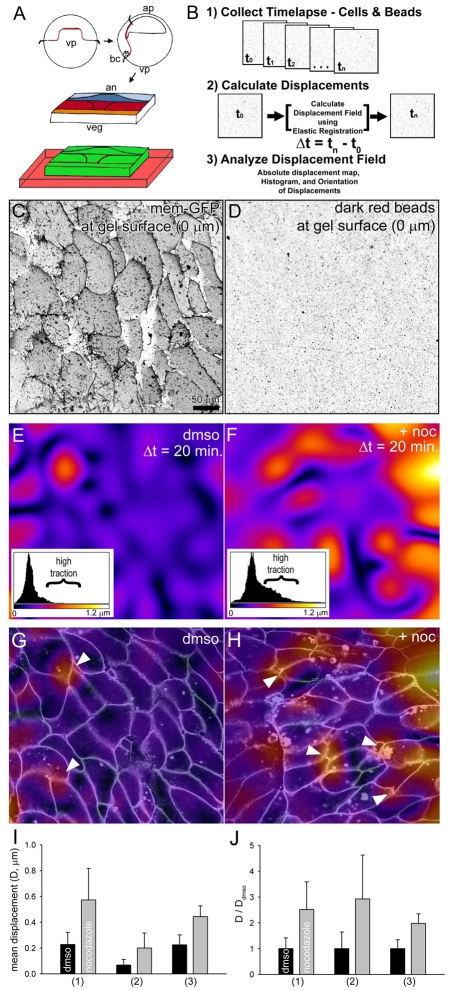
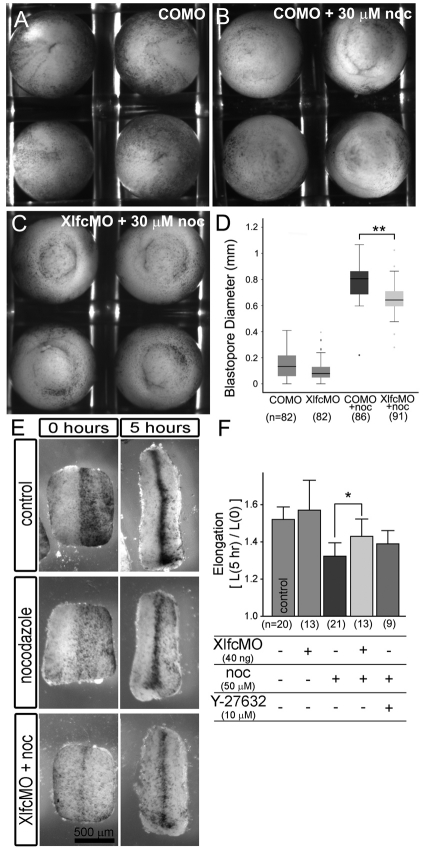
During morphogenesis, forces generated by cells are coordinated and channeled by the viscoelastic properties of the embryo. Microtubules and F-actin are considered to be two of the most important structural elements within living cells accounting for both force production and mechanical stiffness. In this paper, we investigate the contribution of microtubules to the stiffness of converging and extending dorsal tissues in Xenopus laevis embryos using cell biological, biophysical and embryological techniques. Surprisingly, we discovered that depolymerizing microtubules stiffens embryonic tissues by three- to fourfold. We attribute tissue stiffening to Xlfc, a previously identified RhoGEF, which binds microtubules and regulates the actomyosin cytoskeleton. Combining drug treatments and Xlfc activation and knockdown lead us to the conclusion that mechanical properties of tissues such as viscoelasticity can be regulated through RhoGTPase pathways and rule out a direct contribution of microtubules to tissue stiffness in the frog embryo. We can rescue nocodazole-induced stiffening with drugs that reduce actomyosin contractility and can partially rescue morphogenetic defects that affect stiffened embryos. We support these conclusions with a multi-scale analysis of cytoskeletal dynamics, tissue-scale traction and measurements of tissue stiffness to separate the role of microtubules from RhoGEF activation. These findings suggest a re-evaluation of the effects of nocodazole and increased focus on the role of Rho family GTPases as regulators of the mechanical properties of cells and their mechanical interactions with surrounding tissues.
Figures
References
-
- Arganda-Carreras I., Sorzano C. O. S., Marabini R., Carazo J. M., de Solorzano C. O., Kybic J. (2006). Consistent and elastic registration of histological sections using vector-spline regularization. CVAMIA:Computer Vision Approaches to Medical Image Analysis 4241, 85-95
-
- Barentin C., Sawada Y., Rieu J. P. (2006). An iterative method to calculate forces exerted by single cells and multicellular assemblies from the detection of deformations of flexible substrates. Eur. Biophys. J. 35, 328-339 - PubMed
-
- Beningo K. A., Wang Y. L. (2002). Flexible substrata for the detection of cellular traction forces. Trends Cell. Biol. 12, 79-84 - PubMed
-
- Beningo K. A., Lo C. M., Wang Y. L. (2002). Flexible polyacrylamide substrata for the analysis of mechanical interactions at cell-substratum adhesions. Methods Cell Biol. 69, 325-339 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
