

The human papillomavirus type 16 E6 oncoprotein activates mTORC1 signaling and increases protein synthesis
- PMID: 20631133
- PMCID: PMC2937655
- DOI: 10.1128/JVI.00974-10
The human papillomavirus type 16 E6 oncoprotein activates mTORC1 signaling and increases protein synthesis
Abstract
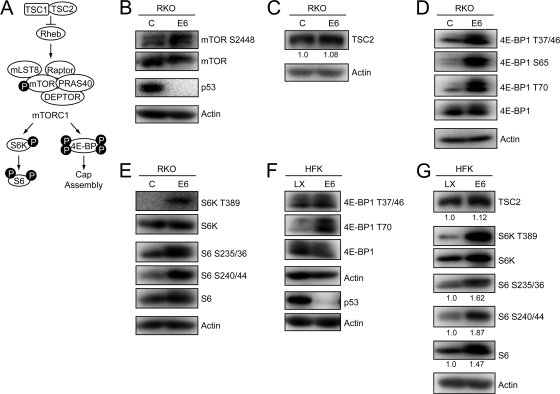
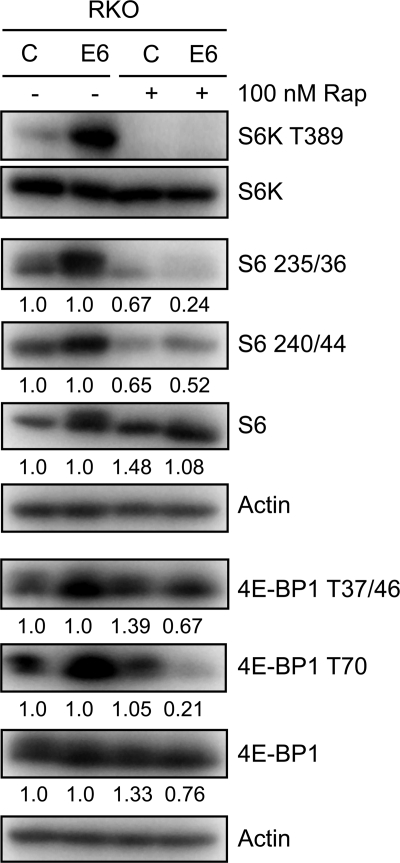
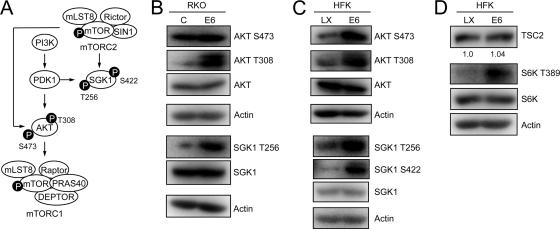
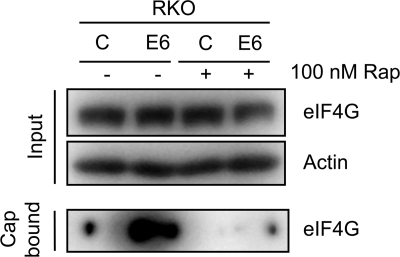
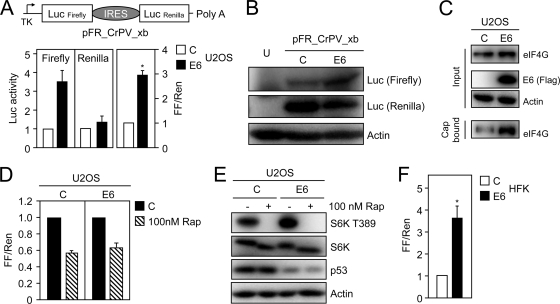
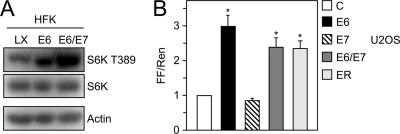
The mammalian target of rapamycin (mTOR) kinase acts as a cellular rheostat that integrates signals from a variety of cellular signal transduction pathways that sense growth factor and nutrient availability as well as intracellular energy status. It was previously reported that the human papillomavirus type 16 (HPV16) E6 oncoprotein may activate the S6 protein kinase (S6K) through binding and E6AP-mediated degradation of the mTOR inhibitor tuberous sclerosis complex 2 (TSC2) (Z. Lu, X. Hu, Y. Li, L. Zheng, Y. Zhou, H. Jiang, T. Ning, Z. Basang, C. Zhang, and Y. Ke, J. Biol. Chem. 279:35664-35670, 2004; L. Zheng, H. Ding, Z. Lu, Y. Li, Y. Pan, T. Ning, and Y. Ke, Genes Cells 13:285-294, 2008). Our results confirmed that HPV16 E6 expression causes an increase in mTORC1 activity through enhanced phosphorylation of mTOR and activation of downstream signaling pathways S6K and eukaryotic initiation factor binding protein 1 (4E-BP1). However, we did not detect a decrease in TSC2 levels in HPV16 E6-expressing cells. We discovered, however, that HPV16 E6 expression causes AKT activation through the upstream kinases PDK1 and mTORC2 under conditions of nutrient deprivation. We show that HPV16 E6 expression causes an increase in protein synthesis by enhancing translation initiation complex assembly at the 5' mRNA cap and an increase in cap-dependent translation. The increase in cap-dependent translation likely results from HPV16 E6-induced AKT/mTORC1 activation, as the assembly of the translation initiation complex and cap-dependent translation are rapamycin sensitive. Lastly, coexpression of the HPV16 E6 and E7 oncoproteins does not affect HPV16 E6-induced activation of mTORC1 and cap-dependent translation. HPV16 E6-mediated activation of mTORC1 signaling and cap-dependent translation may be a mechanism to promote viral replication under conditions of limited nutrient supply in differentiated, HPV oncoprotein-expressing proliferating cells.
Figures
References
-
- Alessi, D. R., S. R. James, C. P. Downes, A. B. Holmes, P. R. Gaffney, C. B. Reese, and P. Cohen. 1997. Characterization of a 3-phosphoinositide-dependent protein kinase which phosphorylates and activates protein kinase Bα. Curr. Biol. 7:261-269. - PubMed
-
- Baker, S. J., S. Markowitz, E. R. Fearon, J. K. Willson, and B. Vogelstein. 1990. Suppression of human colorectal carcinoma cell growth by wild-type p53. Science 249:912-915. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
