

Loss of Topoisomerase I leads to R-loop-mediated transcriptional blocks during ribosomal RNA synthesis
- PMID: 20634320
- PMCID: PMC2904944
- DOI: 10.1101/gad.573310
Loss of Topoisomerase I leads to R-loop-mediated transcriptional blocks during ribosomal RNA synthesis
Abstract
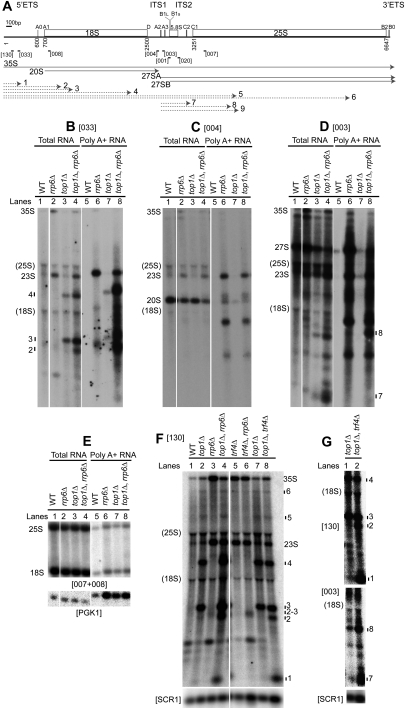
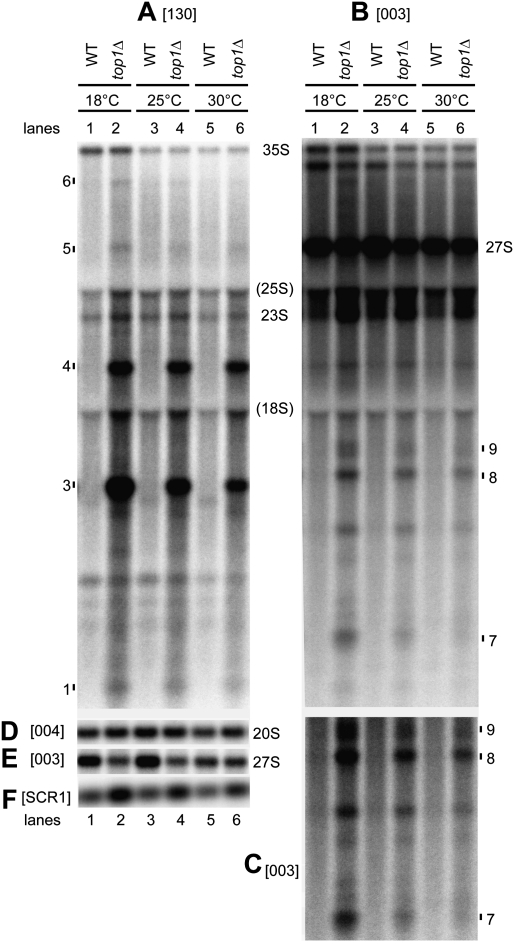
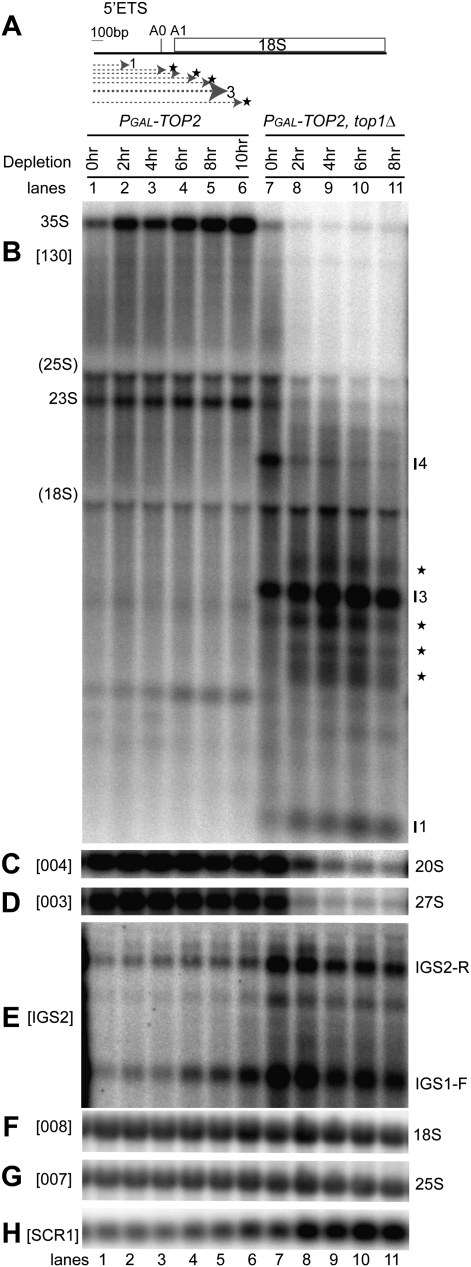
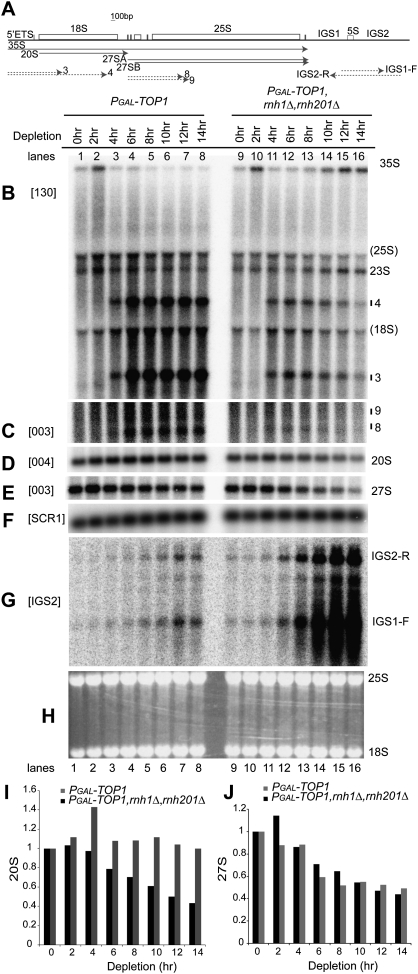
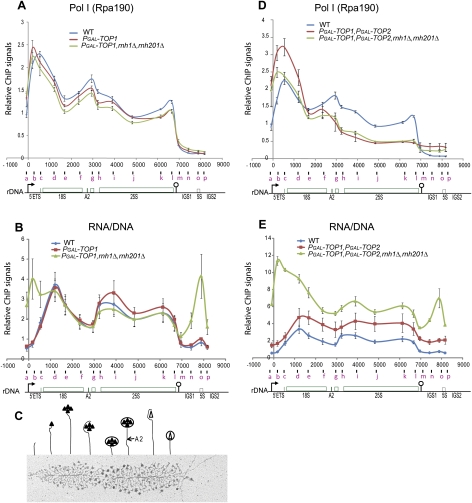
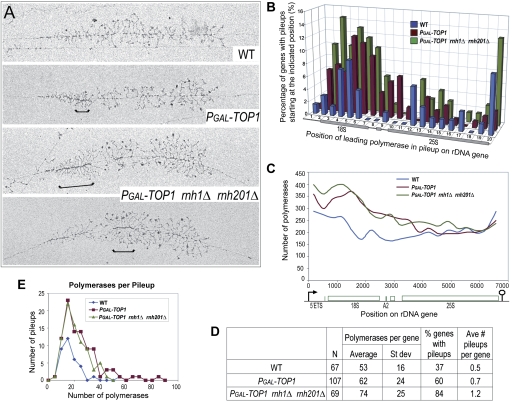
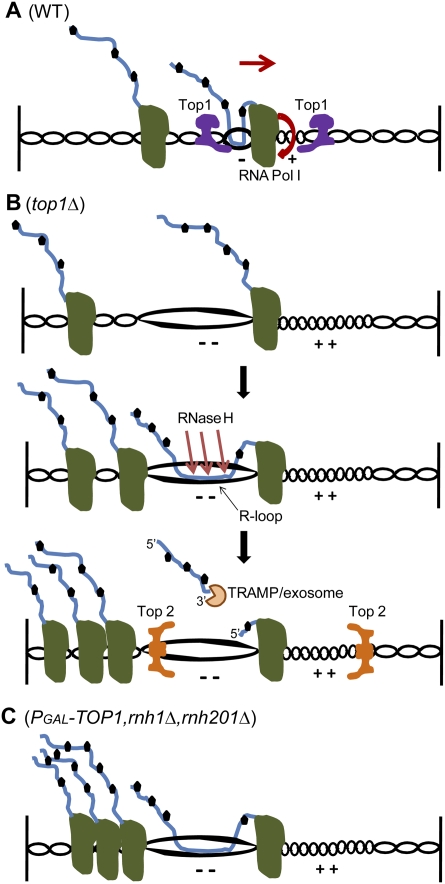
Pre-rRNA transcription by RNA Polymerase I (Pol I) is very robust on active rDNA repeats. Loss of yeast Topoisomerase I (Top1) generated truncated pre-rRNA fragments, which were stabilized in strains lacking TRAMP (Trf4/Trf5-Air1/Air2-Mtr4 polyadenylation complexes) or exosome degradation activities. Loss of both Top1 and Top2 blocked pre-rRNA synthesis, with pre-rRNAs truncated predominately in the 18S 5' region. Positive supercoils in front of Pol I are predicted to slow elongation, while rDNA opening in its wake might cause R-loop formation. Chromatin immunoprecipitation analysis showed substantial levels of RNA/DNA hybrids in the wild type, particularly over the 18S 5' region. The absence of RNase H1 and H2 in cells depleted of Top1 increased the accumulation of RNA/DNA hybrids and reduced pre-rRNA truncation and pre-rRNA synthesis. Hybrid accumulation over the rDNA was greatly exacerbated when Top1, Top2, and RNase H were all absent. Electron microscopy (EM) analysis revealed Pol I pileups in the wild type, particularly over the 18S. Pileups were longer and more frequent in the absence of Top1, and their frequency was exacerbated when RNase H activity was also lacking. We conclude that the loss of Top1 enhances inherent R-loop formation, particularly over the 5' region of the rDNA, imposing persistent transcription blocks when RNase H is limiting.
Figures
References
-
- Aguilera A, Gomez-Gonzalez B 2008. Genome instability: A mechanistic view of its causes and consequences. Nat Rev Genet 9: 204–217 - PubMed
-
- Arud chandran A, Cerritelli S, Narimatsu S, Itaya M, Shin DY, Shimada Y, Crouch RJ 2000. The absence of ribonuclease H1 or H2 alters the sensitivity of Saccharomyces cerevisiae to hydroxyurea, caffeine and ethyl methanesulphonate: Implications for roles of RNases H in DNA replication and repair. Genes Cells 5: 789–802 - PubMed
-
- Boguslawski SJ, Smith DE, Michalak MA, Mickelson KE, Yehle CO, Patterson WL, Carrico RJ 1986. Characterization of monoclonal antibody to DNA.RNA and its application to immunodetection of hybrids. J Immunol Methods 89: 123–130 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous