

Entropy as the driver of chromosome segregation
- PMID: 20634810
- PMCID: PMC3148256
- DOI: 10.1038/nrmicro2391
Entropy as the driver of chromosome segregation
Abstract
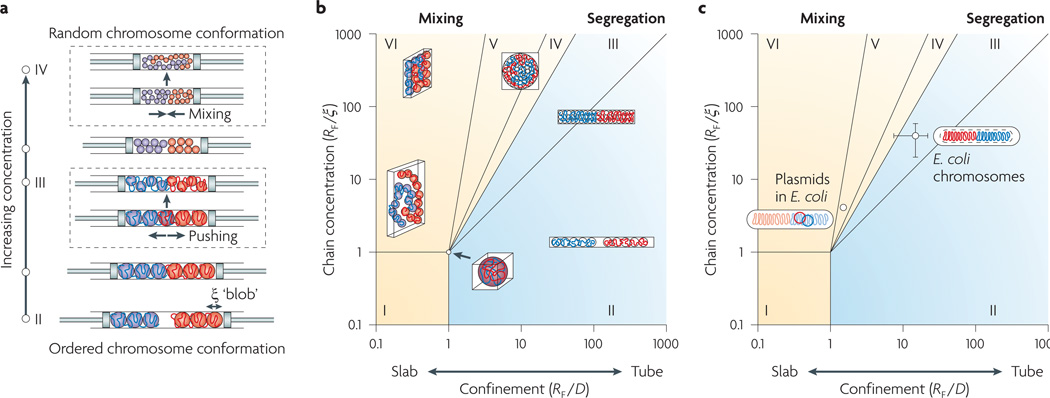
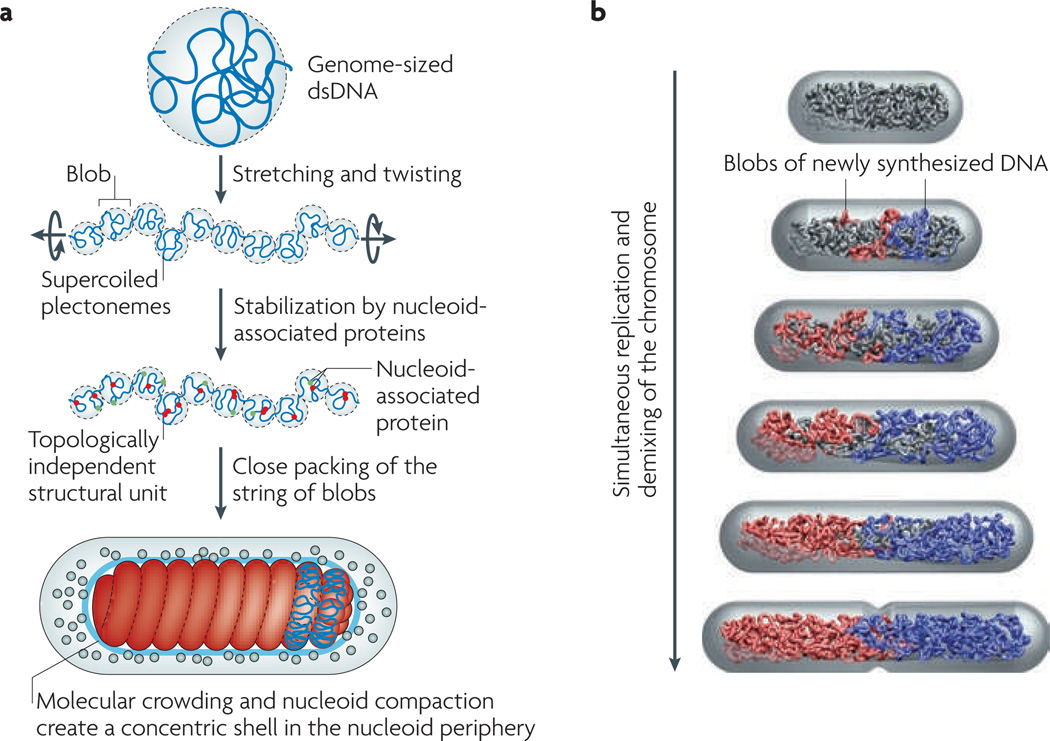
We present a new physical biology approach to understanding the relationship between the organization and segregation of bacterial chromosomes. We posit that replicated Escherichia coli daughter strands will spontaneously demix as a result of entropic forces, despite their strong confinement within the cell; in other words, we propose that entropy can act as a primordial physical force which drives chromosome segregation under the right physical conditions. Furthermore, proteins implicated in the regulation of chromosome structure and segregation may in fact function primarily in supporting such an entropy-driven segregation mechanism by regulating the physical state of chromosomes. We conclude that bacterial chromosome segregation is best understood in terms of spontaneous demixing of daughter strands. Our concept may also have important implications for chromosome segregation in eukaryotes, in which spindle-dependent chromosome movement follows an extended period of sister chromatid demixing and compaction.
Figures
References
-
- Kornberg A, Baker TA. DNA Replication. 2nd edn. New York: Freeman & Company; 1992.
-
- Murray A, Hunt T, editors. The Cell Cycle: an Introduction. New York: Oxford Univ. Press; 1993.
-
- Wang JC. DNA topoisomerases. Annu. Rev. Biochem. 1996;65:635–692. - PubMed
-
- Gruber S, Errington J. Recruitment of condensin to replication origin regions by ParB/SpoOJ promotes chromosome segregation in B. subtilis. Cell. 2009;137:685–696. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
