

Curcumin increases the pathogenicity of Salmonella enterica serovar Typhimurium in murine model
- PMID: 20634977
- PMCID: PMC2901387
- DOI: 10.1371/journal.pone.0011511
Curcumin increases the pathogenicity of Salmonella enterica serovar Typhimurium in murine model
Abstract
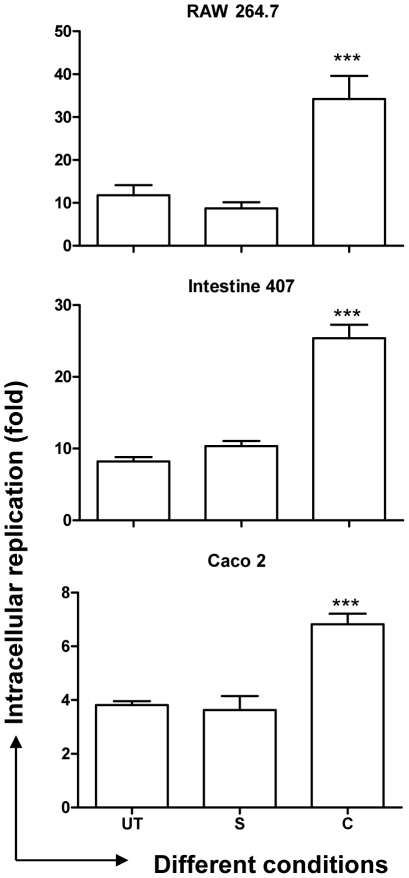
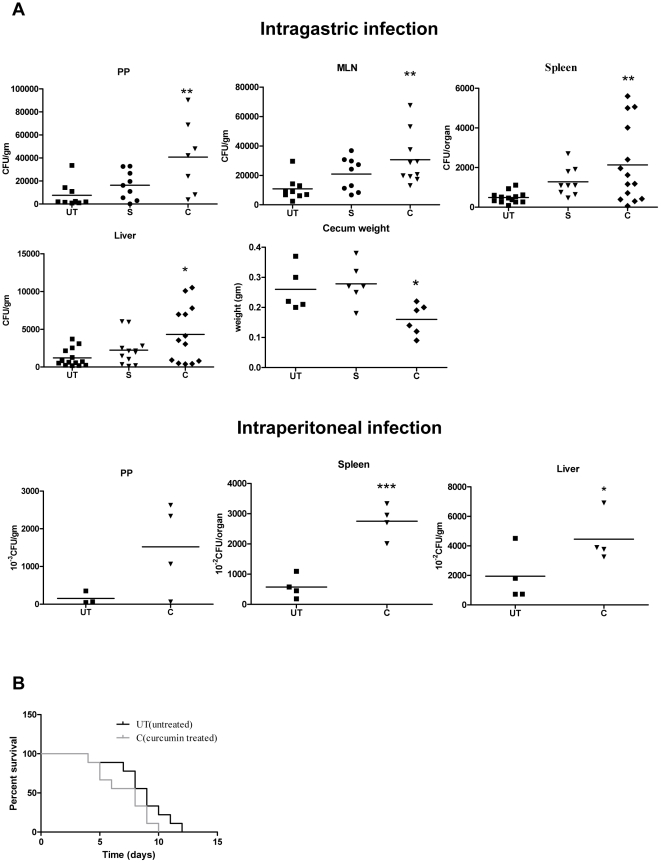
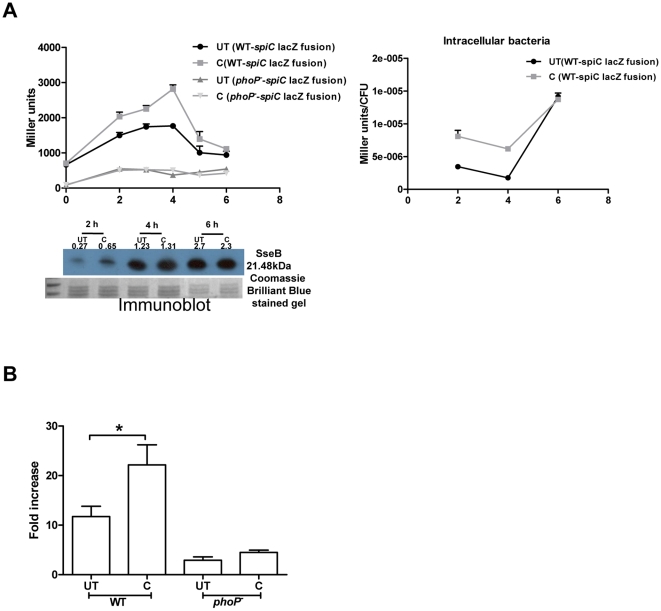
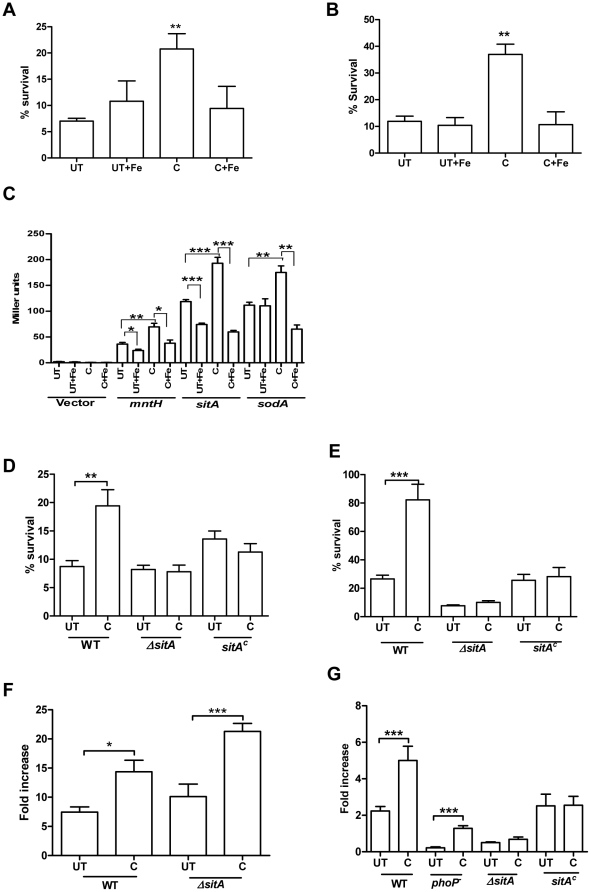
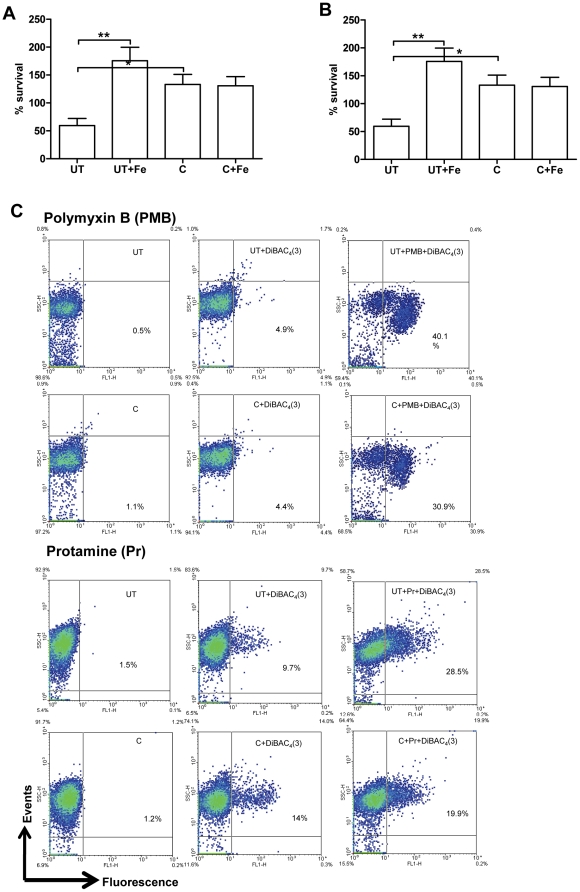
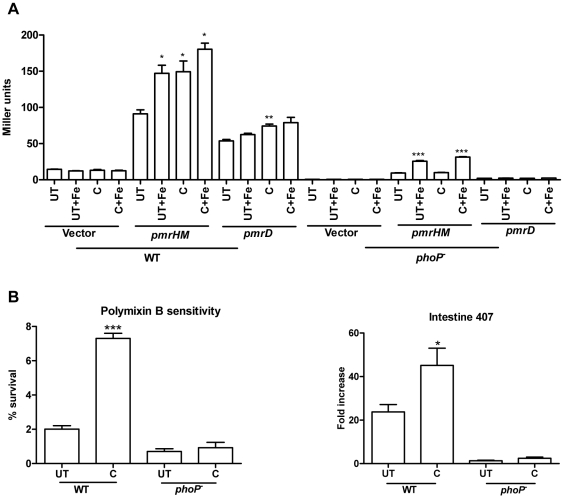
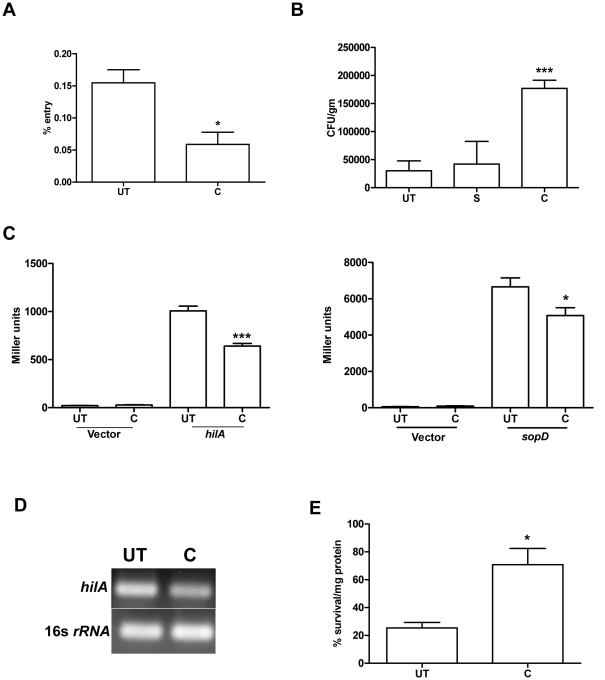
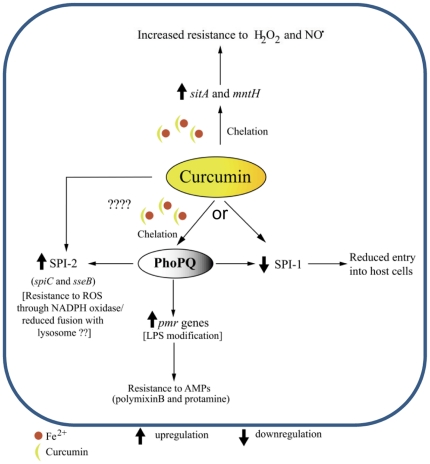
Curcumin has gained immense importance for its vast therapeutic and prophylactic applications. Contrary to this, our study reveals that it regulates the defense pathways of Salmonella enterica serovar Typhimurium (S. Typhimurium) to enhance its pathogenicity. In a murine model of typhoid fever, we observed higher bacterial load in Peyer's patches, mesenteric lymph node, spleen and liver, when infected with curcumin-treated Salmonella. Curcumin increased the resistance of S. Typhimurium against antimicrobial agents like antimicrobial peptides, reactive oxygen and nitrogen species. This increased tolerance might be attributed to the up-regulation of genes involved in resistance against antimicrobial peptides--pmrD and pmrHFIJKLM and genes with antioxidant function--mntH, sodA and sitA. We implicate that iron chelation property of curcumin have a role in regulating mntH and sitA. Interestingly, we see that the curcumin-mediated modulation of pmr genes is through the PhoPQ regulatory system. Curcumin downregulates SPI1 genes, required for entry into epithelial cells and upregulates SPI2 genes required to intracellular survival. Since it is known that the SPI1 and SPI2 system can be regulated by the PhoPQ system, this common regulator could explain curcumin's mode of action. This data urges us to rethink the indiscriminate use of curcumin especially during Salmonella outbreaks.
Conflict of interest statement
Figures
References
-
- Sharma RA, Gescher AJ, Steward WP. Curcumin: the story so far. Eur J Cancer. 2005;41:1955–1968. - PubMed
-
- Aggarwal BB, Sundaram C, Malani N, Ichikawa H. Curcumin: the Indian solid gold. Adv Exp Med Biol. 2007;595:1–75. - PubMed
-
- Dahl TA, McGowan WM, Shand MA, Srinivasan VS. Photokilling of bacteria by the natural dye curcumin. Arch Microbiol. 1989;151:183–185. - PubMed
-
- Rai D, Singh JK, Roy N, Panda D. Curcumin inhibits FtsZ assembly: an attractive mechanism for its antibacterial activity. Biochem J. 2008;410:147–155. - PubMed
-
- Han C, Wang L, Yu K, Chen L, Hu L, et al. Biochemical characterization and inhibitor discovery of shikimate dehydrogenase from Helicobacter pylori. Febs J. 2006;273:4682–4692. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
