

doi: 10.1016/j.ijbiomac.2010.07.001.
Epub 2010 Jul 14.
Role of tyrosine 33 residue for the stabilization of the tetrameric structure of human cytidine deaminase
Affiliations
- PMID: 20637228
- PMCID: PMC3020594
- DOI: 10.1016/j.ijbiomac.2010.07.001
Item in Clipboard
Role of tyrosine 33 residue for the stabilization of the tetrameric structure of human cytidine deaminase
Int J Biol Macromol.
.
Erratum in
- Int J Biol Macromol. 2011 Oct 1;49(3):439
Abstract
In the present work the effect of a mutation on tyrosine 33 residue (Y33G) of human cytidine deaminase (CDA) was investigated with regard to protein solubility and specific activity. Osmolytes and CDA ligands were used to increase the yield and the specific activity of the protein. The mutant enzyme was purified and subjected to a kinetic characterization and to stability studies. These investigations reinforced the hypothesis that in human CDA the side chain of Y33 is involved in intersubunit interactions with four glutamate residues (E108) forming a double latch that connects each of the two pairs of monomers of the tetrameric CDA.
Copyright © 2010 Elsevier B.V. All rights reserved.
Figures
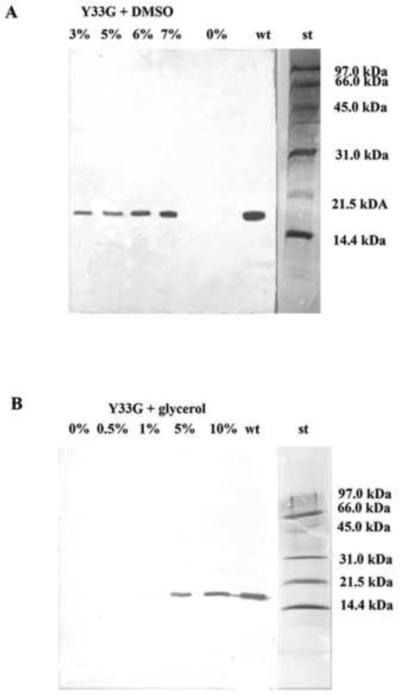
Western blot analysis to evaluate the expression of the mutant CDA Y33G in presence of low molecular weight compounds in the culture medium. A) DMSO 0%, 3%, 5%, 6%, 7%; B) Glycerol 0%, 0.5%, 1%, 5%, 10%, wt: wild-type CDA (positive control); st: Bio-Rad low molecular weight standard (97.4 kDa, phosphorylase b; 66.2 kDa, bovine serum albumin; 45.0 kDa, ovalbumin; 31.0 kDa, carbonic anhydrase; 21.5 kDa, soybean trypsin inhibitor; 14.4 kDa, lysozyme).
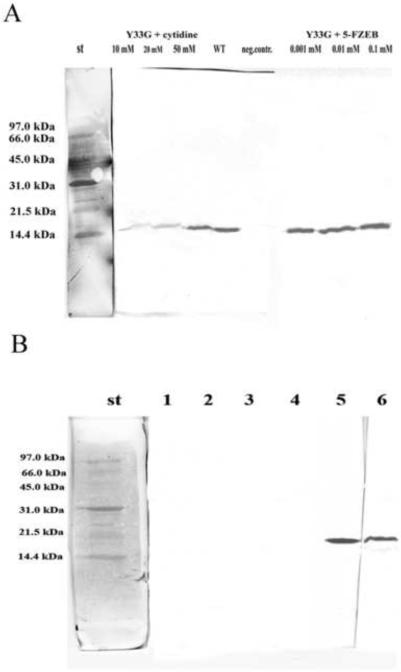
A) Western blot analysis to evaluate the expression of the mutant CDA Y33G in presence of specific ligands of cytidine deaminase in the culture medium. wt: wild-type CDA (positive control); neg. contr: no CDA ligands; st: Bio-Rad low molecular weight standard. B) Western blot analysis to evaluate the presence of inclusion bodies in the crude extracts of Y33G. 1: no glycerol in the culture medium. E. coli cells were lysed and after centrifugation the supernatant was resuspended in buffer A without glycerol and analyzed; 2 and 4: no glycerol in the culture medium. E. coli cells were lysed and after centrifugation the pellet was resuspended in 8M urea and analyzed; 3: no glycerol in the culture medium. E. coli cells were lysed and after centrifugation the supernatant was resuspended in buffer A with10 % glycerol and analyzed; 5: 10% glycerol in the culture medium. E. coli cells were lysed and after centrifugation the supernatant was resuspended in buffer A with 10 % glycerol and analyzed; 6: 10% glycerol in the culture medium. E. coli cells were lysed and after centrifugation the pellet was resuspended in 8M urea and analyzed. St: Bio-Rad low molecular weight standard
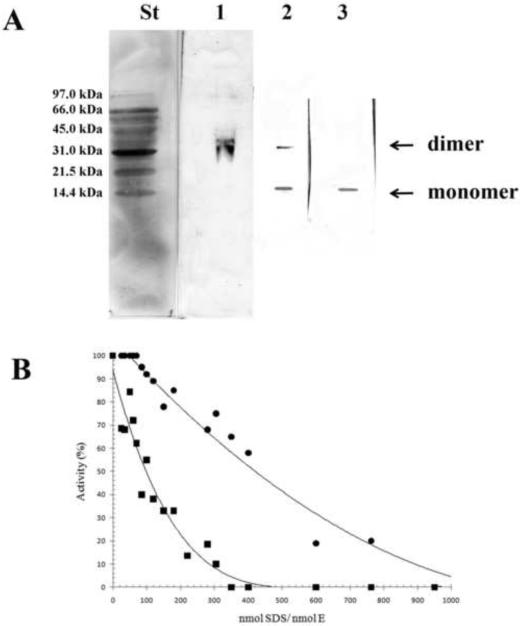
A) 15% PAGE followed by western blot analysis of Y33G at different SDS concentrations. Each lane represents separate gels with different concentrations of the denaturing agent. The SDS/enzyme molar ratios in the individual lanes were as follows: lane 1, 70; lane 2, 120; lane 3, 300. St, Bio-Rad low molecular weight standard. B) Percentage of enzymatic activity of wild-type CDA (●) andY33G mutant CDA (■) after incubations with SDS concentrations ranging from 0.35 to 1.73 mM in order to give different SDS/enzyme molar ratios. The data of wild type CDA are from [20].
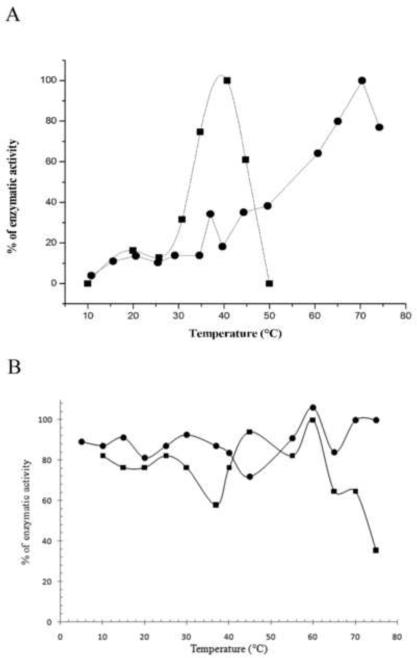
A) Temperature effect on the activity of wild-type CDA (●) and Y33G mutant enzyme (■) obtained by plotting the percentage of enzymatic activity at temperatures ranging from 10 to 80°C. B) Effect of temperature on enzyme stability of wild-type CDA (●) and Y33G mutant CDA(■).
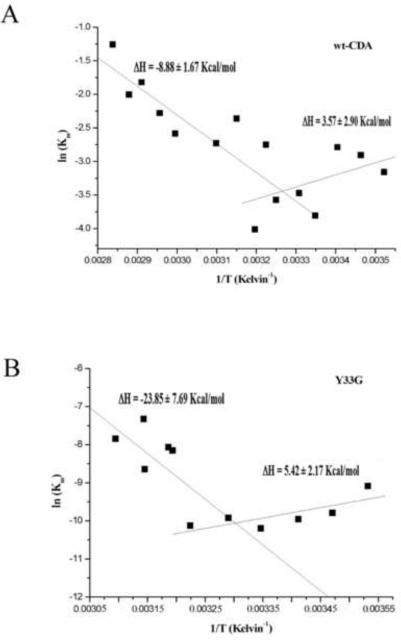
Van't Hoff plot for wild-type CDA (A) and for Y33G mutant CDA (B).
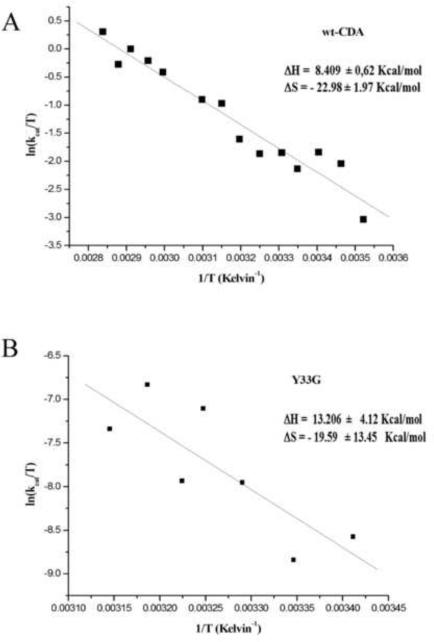
Eyring plot for wild-type CDA (A) and Y33G mutant CDA (B).
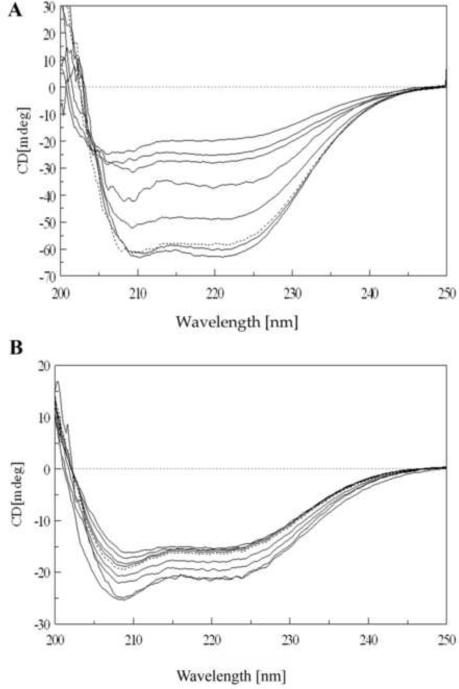
A) CD spectra of Y33G temperature unfolding experiment: protein concentration was 5 μM in 20 mM phosphate buffer pH 7.5 in the presence of 1 mM DTT and 10% glycerol. Temperatures are from bottom to top (at 222 nm) 5, 40, 50, 60, 70, 80 and 90° C. Dotted line is the spectrum of the protein sample refolded by cooling the solution from 90° C to 5° C. Spectra obtained at 10, 20 and 30°C are omitted for the sake of clarity (see text). B) CD spectra of Y33G SDS unfolding experiment: protein concentration was 2 μM in 20 mM phosphate buffer pH 7.5 in the presence of 1 mM DTT and 10% glycerol. Temperature was 20°C. SDS concentrations are from top to bottom (at 210 nm) 0.035, 0.69, 1.04, 1.40, 1.70, 3.5, 5.2, 6.9 mM. Dotted line is the spectrum of protein sample refolded after extensive dialysis against buffer A. Spectra obtained at SDS concentrations of 0.069, 0.17, 0.35, 0.52 mM are omitted for the sake of clarity (see text).
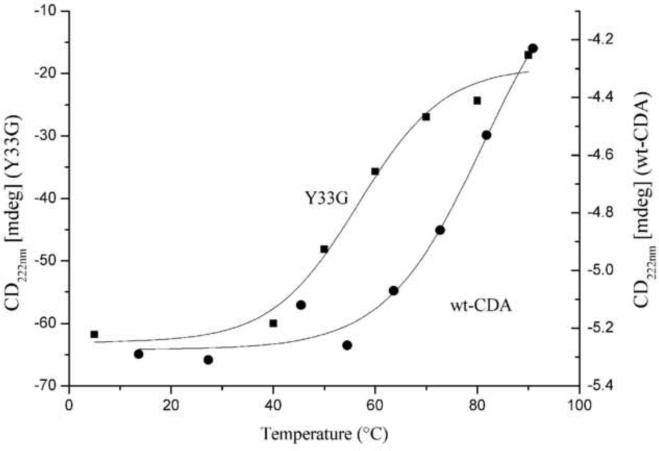
Thermal unfolding analysis of the CD signal of the wild type (●) and Y33G (■) CDAs (performed as described in Materials and Methods) plotted against the temperature (from 5 to 90°C). The difference of the Y axis scale is due to a different protein concentration and cell length used on the two series of experiments.
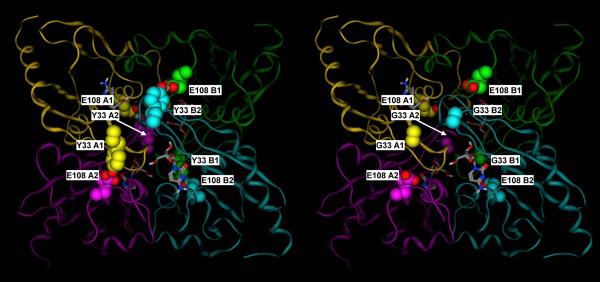
A) Crystal structure of the mouse cytidine deaminase in complex with cytidine (2FR6, (Teh et al., 2006). B) Corresponding molecular model of the Y33 mutant. Y33 of subunits A1, A2, B1 and B2 form hydrogen bonds with E108 of subunits A2, A1, B2 and B1, respectively, thus double latching each of the two monomer pairs corresponding to the catalytic site and the C-terminal broken site in the E. coli dimeric enzyme (A1–A2 and B1–B2). Subunit A1, A2, B1 and B2 are in yellow, purple, green and cyan, respectively. Y33 and E108 residues are shown in space filling representation, with the carbon atoms colored by subunit, and the heteroatoms colored by element. Cytidine is shown in sticks representation, with the atoms colored by element.
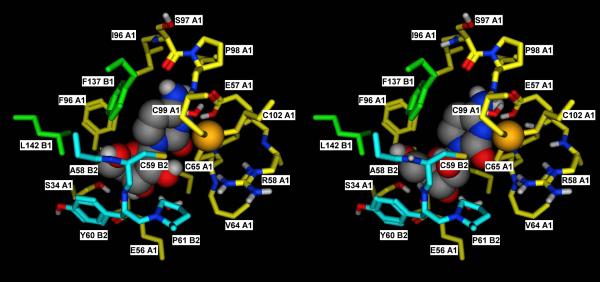
A) Detailed view of the catalytic site of subunit A1 in the crystal structure of the mouse cytidine deaminase in complex with cytidine (2FR6, (Teh et al., 2006)). B) Corresponding docking complex obtained in silico for Ara-C. The catalytic importance of the tetrameric structure derives from the fact that residues of subunits B1 and B2 contribute to the formation of the catalytic site. The docking complex obtained for Ara-C suggests that the orientation that the 2'-OH group assumes in this compound may exert some steric hindrance with the residues of subunit B2, thus adding a destabilizing element for the quaternary structure. In the Y33 mutant, this is probably sufficient to disrupt the already weak tetramer, thus explaining its inability to catalyze the deamination of Ara-C. Subunit A1, B1 and B2 are in yellow, green and cyan, respectively. Residues lining the binding pocket are shown in sticks representation, with the carbon atoms colored by subunit, and the heteroatoms colored by element. Cytidine and Ara-C are shown in space filling representation, with the atoms colored by element.
References
-
- Vincenzetti S, Quadrini B, Mariani P, De Sanctis G, Cammertoni N, Polzonetti V, Pucciarelli S, Natalini P, Vita A. Modulation of human cytidine deaminase by specific aminoacids involved in the intersubunit interactions. Proteins. 2008;70:144–156. - PubMed
-
- Costanzi S, Vincenzetti S, Cristalli G, Vita A. Human cytidine deaminase: a three-dimensional model of a tetrameric metallo-enzyme inferred from a crystal structure of a distantly related dimeric homologue. J. Mol. Graph. Model. 2006;25:10–16. - PubMed
-
- Betts L, Xiang S, Short SA, Wolfenden R, Carter CW. Cytidine deaminase. The 2.3 Ǻ crystal structure of an enzyme: transition-state analog complex. J. Mol. Biol. 1994;235:635–656. - PubMed
-
- Johansson E, Mejlhede N, Neuhard J, Larsen S. Crystal structure of the tetrameric cytidine deaminase from Bacillus subtilis at 2.0 Å resolution. Biochemistry. 2002;41:2563–2570. - PubMed
-
- Vincenzetti S, Cambi A, Maury G, Bertorelle F, Gaubert G, Neuhard J, Natalini P, Salvatori D, De Sanctis G, Vita A. Possible role of two phenylalanine residues in the active site of human cytidine deaminase. Protein Eng. 2000;13:791–799. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
