

The paralogous MarR/DUF24-family repressors YodB and CatR control expression of the catechol dioxygenase CatE in Bacillus subtilis
- PMID: 20639328
- PMCID: PMC2937424
- DOI: 10.1128/JB.00409-10
The paralogous MarR/DUF24-family repressors YodB and CatR control expression of the catechol dioxygenase CatE in Bacillus subtilis
Abstract
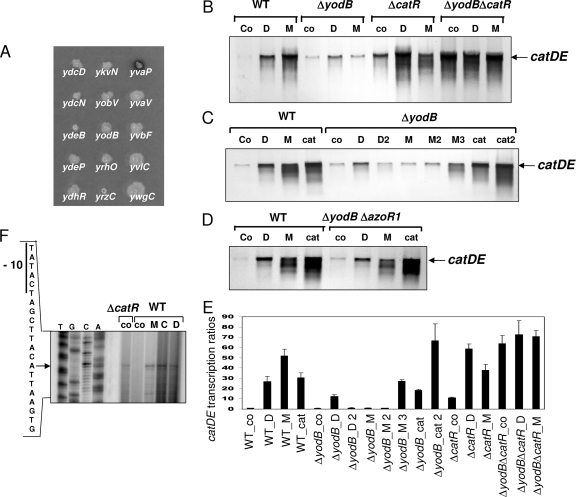
The redox-sensing MarR/DUF24-type repressor YodB controls expression of the azoreductase AzoR1 and the nitroreductase YodC that are involved in detoxification of quinones and diamide in Bacillus subtilis. In the present paper, we identified YodB and its paralog YvaP (CatR) as repressors of the yfiDE (catDE) operon encoding a catechol-2,3-dioxygenase that also contributes to quinone resistance. Inactivation of both CatR and YodB is required for full derepression of catDE transcription. DNA-binding assays and promoter mutagenesis studies showed that CatR protects two inverted repeats with the consensus sequence TTAC-N(5)-GTAA overlapping the -35 promoter region (BS1) and the transcriptional start site (TSS) (BS2). The BS1 operator was required for binding of YodB in vitro. CatR and YodB share the conserved N-terminal Cys residue, which is required for redox sensing of CatR in vivo as shown by Cys-to-Ser mutagenesis. Our data suggest that CatR is modified by intermolecular disulfide formation in response to diamide and quinones in vitro and in vivo. Redox regulation of CatR occurs independently of YodB, and no protein interaction was detected between CatR and YodB in vivo using protein cross-linking and mass spectrometry.
Figures








References
-
- Antelmann, H., M. Hecker, and P. Zuber. 2008. Proteomic signatures uncover thiol-specific electrophile resistance mechanisms in Bacillus subtilis. Expert Rev. Proteomics 5:77-90. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
