

doi: 10.1128/JB.00371-10.
Epub 2010 Jul 16.
Aromatic acid metabolites of Escherichia coli K-12 can induce the marRAB operon
Affiliations
- PMID: 20639340
- PMCID: PMC2937402
- DOI: 10.1128/JB.00371-10
Item in Clipboard
Aromatic acid metabolites of Escherichia coli K-12 can induce the marRAB operon
J Bacteriol.
2010 Sep.
Abstract
MarR is a key regulator of the marRAB operon involved in antibiotic resistance and solvent stress tolerance in Escherichia coli. We show that two metabolic intermediates, 2,3-dihydroxybenzoate and anthranilate, involved in enterobactin and tryptophan biosynthesis, respectively, can activate marRAB transcription. We also found that a third intermediate involved in ubiquinone biosynthesis, 4-hydroxybenzoate, activates marRAB transcription in the absence of TolC. Of the three, however, only 2,3-dihydroxybenzoate directly binds MarR and affects its activity.
Figures
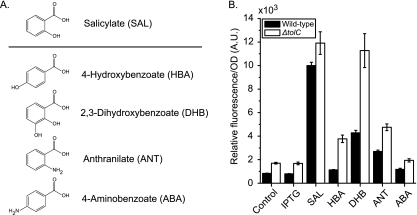
Activation of the marRAB operon by aromatic acid metabolites in the superpathway of chorismate. (A) Chemical structure of metabolites selected based on their similarity to salicylate. (B) Observed transcriptional activation of the marR′-yfp promoter fusion in the presence of 5 mM concentrations of indicated inducers in wild-type (CR700) and ΔtolC mutant (CR703) backgrounds. Salicylate and IPTG served as positive and negative controls for activation, respectively. Cells were grown overnight in MOPS (morpholinepropanesulfonic acid) minimal medium (20 mM glucose, 0.2% Casamino Acids, pH 7.2) and subcultured 1:200 in fresh medium. Following dilution, 450 μl of culture was transferred to deep 96-well plates and grown at 37°C with aeration at 1,000 rpm to an OD of 0.5. At this time, 100 μl of medium containing dissolved inducer was added. Growth was continued for an additional 2 h prior to fluorescence and optical density measurements made with a Tecan Safire2 microplate reader. An expanded description of the experimental procedures is provided in the supplementary material. A.U., arbitrary units.
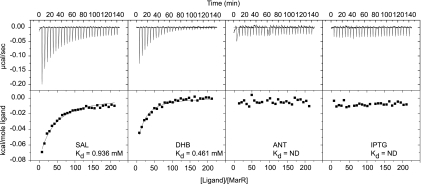
MarR binding of aromatic acid inducers as determined by isothermal titration calorimetry (ITC). Experiments were conducted using a VP-ITC calorimeter (MicroCal) with 1.4 ml MarR at 10 μM and ligands at a concentration of 10 mM, both in Tris-buffered saline (50 mM Tris, 150 mM NaCl, pH 7.4). In the cases of ANT and HBA, higher concentrations of ligand were also tested though again no binding was observed (data not shown). Titration reactions were performed with 28 injections, all 10 μl in volume, with constant stirring at 300 rpm at 25°C. Data acquisition and binding coefficients were determined with the Origin-based MicroCal analysis software.

MarR DNA binding activity in the presence of aromatic acid metabolites as determined by electromobility shift assays. Binding reaction mixtures consisted of 20 ng of purified MarR and 5 ng of a radiolabeled, 150-bp fragment of the marRAB promoter containing two MarR operator sites. The ligands salicylate (SAL), 2,3-dihydroxybenzoate (DHB), anthranilate (ANT), and IPTG were supplied at increasing concentrations of 2.5 mM, 5 mM, and 10 mM to the binding reaction mixtures. Reactions were displayed on 5% acrylamide, 0.5× Tris-borate-EDTA (TBE)-buffered gels. Loss of MarR DNA binding activity was monitored by the emergence of free DNA in the presence of these ligands. Salicylate and IPTG served as positive and negative controls, respectively. A detailed description of the methods is provided in the supplemental material.
References
-
- Alekshun, M. N., S. B. Levy, T. R. Mealy, B. A. Seaton, and J. F. Head. 2001. The crystal structure of MarR, a regulator of multiple antibiotic resistance, at 2.3 Å resolution. Nat. Struct. Biol. 8:710-714. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
