

The reorganization and reactivation of hippocampal maps predict spatial memory performance
- PMID: 20639874
- PMCID: PMC2923061
- DOI: 10.1038/nn.2599
The reorganization and reactivation of hippocampal maps predict spatial memory performance
Abstract
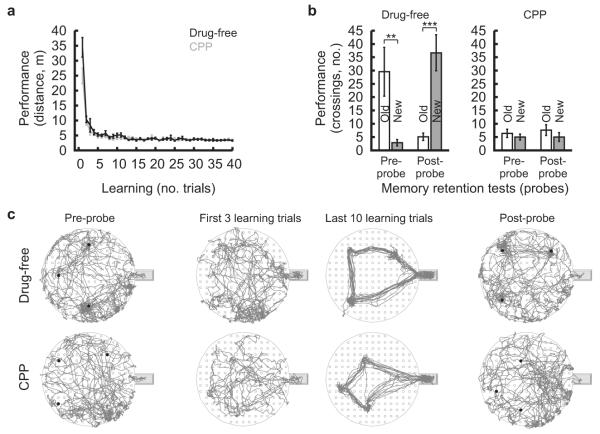
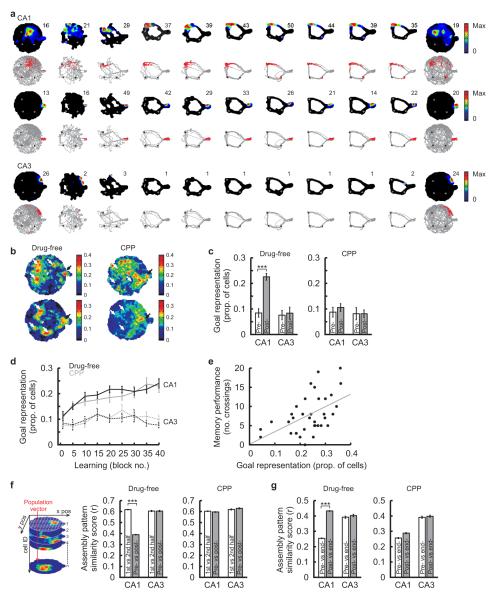
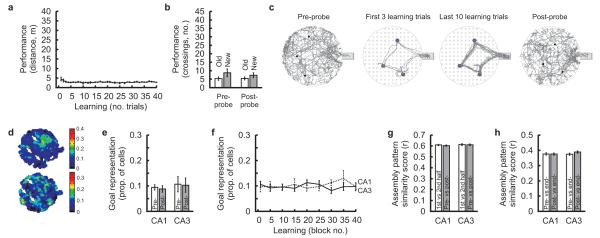
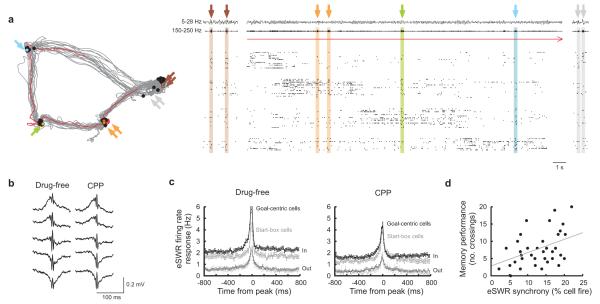
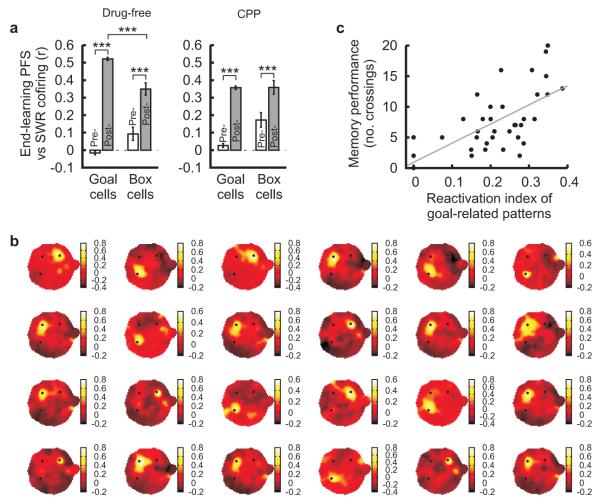
The hippocampus is an important brain circuit for spatial memory and the spatially selective spiking of hippocampal neuronal assemblies is thought to provide a mnemonic representation of space. We found that remembering newly learnt goal locations required NMDA receptor-dependent stabilization and enhanced reactivation of goal-related hippocampal assemblies. During spatial learning, place-related firing patterns in the CA1, but not CA3, region of the rat hippocampus were reorganized to represent new goal locations. Such reorganization did not occur when goals were marked by visual cues. The stabilization and successful retrieval of these newly acquired CA1 representations of behaviorally relevant places was NMDAR dependent and necessary for subsequent memory retention performance. Goal-related assembly patterns associated with sharp wave/ripple network oscillations, during both learning and subsequent rest periods, predicted memory performance. Together, these results suggest that the reorganization and reactivation of assembly firing patterns in the hippocampus represent the formation and expression of new spatial memory traces.
Figures
Comment in
-
Who moved my cheese (again)?Nat Neurosci. 2010 Aug;13(8):911-2. doi: 10.1038/nn0810-911. Nat Neurosci. 2010. PMID: 20661265
References
-
- O’Keefe J, Nadel L. The hippocampus as a cognitive map. Oxford University Press; Oxford,Clarendon: 1978.
-
- Squire LR. Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans. Psychol. Rev. 1992;99:195–231. - PubMed
-
- Morris RG. Elements of a neurobiological theory of hippocampal function: the role of synaptic plasticity, synaptic tagging and schemas. Eur. J. Neurosci. 2006;23:2829–2846. - PubMed
-
- Riedel G, et al. Reversible neural inactivation reveals hippocampal participation in several memory processes. Nat. Neurosci. 1999;2:898–905. - PubMed
-
- Hebb DO. The organization of behavior. Wiley; New York: 1949.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
