

The ATP-sensitive K(+)-channel (K(ATP)) controls early left-right patterning in Xenopus and chick embryos
- PMID: 20643119
- PMCID: PMC2937067
- DOI: 10.1016/j.ydbio.2010.07.011
The ATP-sensitive K(+)-channel (K(ATP)) controls early left-right patterning in Xenopus and chick embryos
Abstract
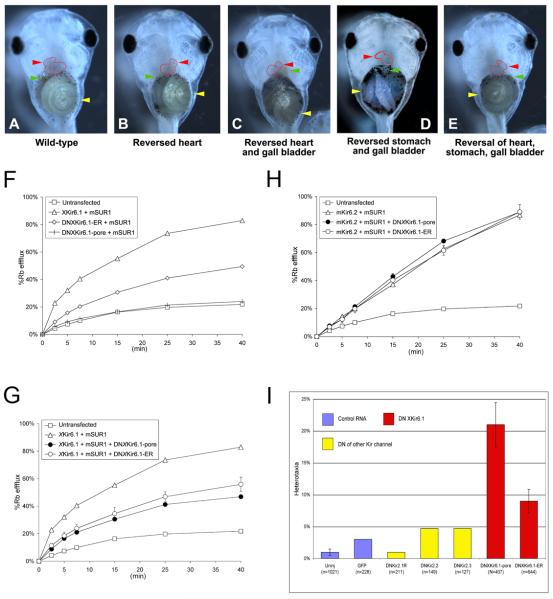
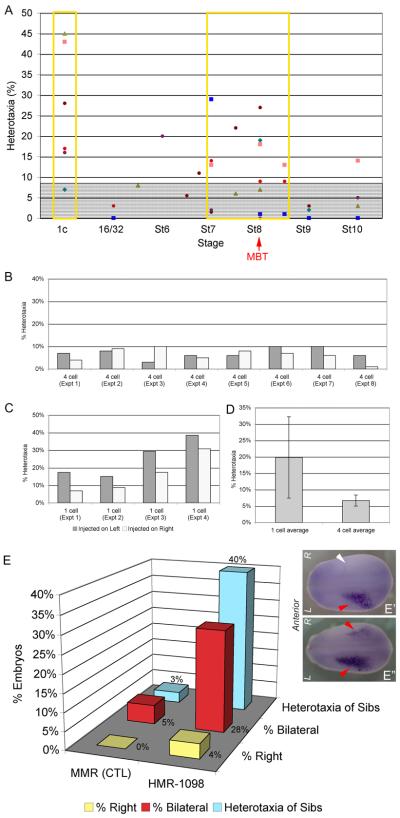
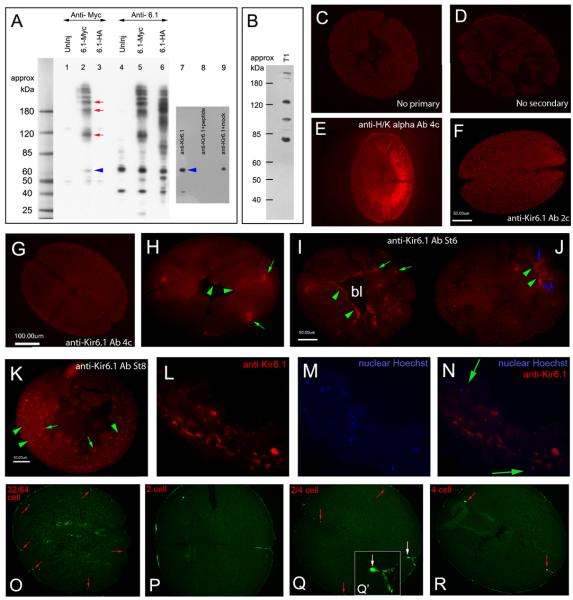
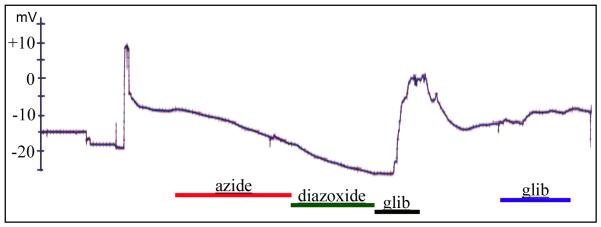
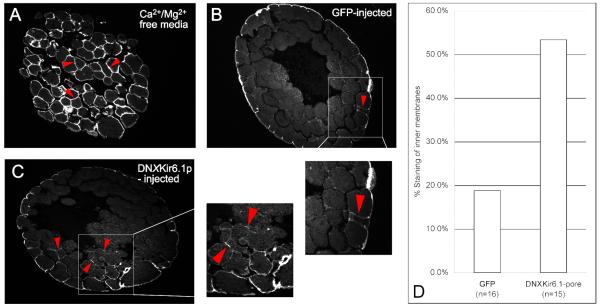
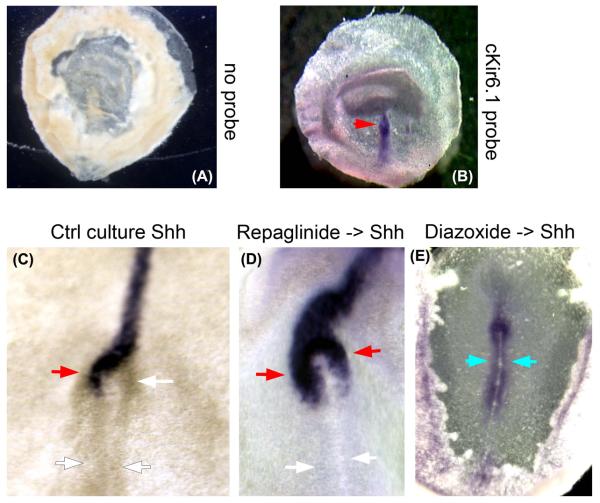
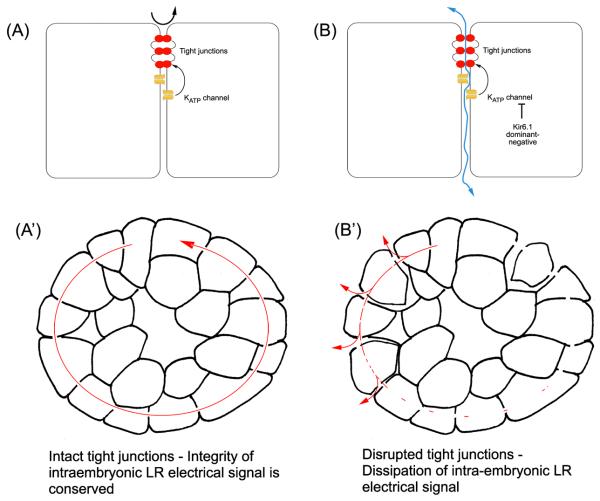
Consistent left-right asymmetry requires specific ion currents. We characterize a novel laterality determinant in Xenopus laevis: the ATP-sensitive K(+)-channel (K(ATP)). Expression of specific dominant-negative mutants of the Xenopus Kir6.1 pore subunit of the K(ATP) channel induced randomization of asymmetric organ positioning. Spatio-temporally controlled loss-of-function experiments revealed that the K(ATP) channel functions asymmetrically in LR patterning during very early cleavage stages, and also symmetrically during the early blastula stages, a period when heretofore largely unknown events transmit LR patterning cues. Blocking K(ATP) channel activity randomizes the expression of the left-sided transcription of Nodal. Immunofluorescence analysis revealed that XKir6.1 is localized to basal membranes on the blastocoel roof and cell-cell junctions. A tight junction integrity assay showed that K(ATP) channels are required for proper tight junction function in early Xenopus embryos. We also present evidence that this function may be conserved to the chick, as inhibition of K(ATP) in the primitive streak of chick embryos randomizes the expression of the left-sided gene Sonic hedgehog. We propose a model by which K(ATP) channels control LR patterning via regulation of tight junctions.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
H,K-ATPase protein localization and Kir4.1 function reveal concordance of three axes during early determination of left-right asymmetry.Mech Dev. 2008 Mar-Apr;125(3-4):353-72. doi: 10.1016/j.mod.2007.10.011. Epub 2007 Nov 4. Mech Dev. 2008. PMID: 18160269 Free PMC article.
-
Gap junction-mediated transfer of left-right patterning signals in the early chick blastoderm is upstream of Shh asymmetry in the node.Development. 1999 Nov;126(21):4703-14. doi: 10.1242/dev.126.21.4703. Development. 1999. PMID: 10518488
-
Serotonin signaling is a very early step in patterning of the left-right axis in chick and frog embryos.Curr Biol. 2005 May 10;15(9):794-803. doi: 10.1016/j.cub.2005.03.044. Curr Biol. 2005. PMID: 15886096
-
K(ATP) channel action in vascular tone regulation: from genetics to diseases.Sheng Li Xue Bao. 2012 Feb 25;64(1):1-13. Sheng Li Xue Bao. 2012. PMID: 22348955 Free PMC article. Review.
-
Conserved roles for cytoskeletal components in determining laterality.Integr Biol (Camb). 2016 Mar 14;8(3):267-86. doi: 10.1039/c5ib00281h. Integr Biol (Camb). 2016. PMID: 26928161 Free PMC article. Review.
Cited by
-
Gap junctional signaling in pattern regulation: Physiological network connectivity instructs growth and form.Dev Neurobiol. 2017 May;77(5):643-673. doi: 10.1002/dneu.22405. Epub 2016 Jun 24. Dev Neurobiol. 2017. PMID: 27265625 Free PMC article. Review.
-
Electrochemical patterns during Drosophila oogenesis: ion-transport mechanisms generate stage-specific gradients of pH and membrane potential in the follicle-cell epithelium.BMC Dev Biol. 2019 Jun 21;19(1):12. doi: 10.1186/s12861-019-0192-x. BMC Dev Biol. 2019. PMID: 31226923 Free PMC article.
-
Low frequency vibrations disrupt left-right patterning in the Xenopus embryo.PLoS One. 2011;6(8):e23306. doi: 10.1371/journal.pone.0023306. Epub 2011 Aug 3. PLoS One. 2011. PMID: 21826245 Free PMC article.
-
Information integration during bioelectric regulation of morphogenesis of the embryonic frog brain.iScience. 2023 Nov 4;26(12):108398. doi: 10.1016/j.isci.2023.108398. eCollection 2023 Dec 15. iScience. 2023. PMID: 38034358 Free PMC article.
-
Bioelectric gene and reaction networks: computational modelling of genetic, biochemical and bioelectrical dynamics in pattern regulation.J R Soc Interface. 2017 Sep;14(134):20170425. doi: 10.1098/rsif.2017.0425. J R Soc Interface. 2017. PMID: 28954851 Free PMC article.
References
-
- Adams DS, Levin M. Strategies and techniques for investigation of biophysical signals in patterning. In: Whitman M, Sater AK, editors. Analysis of Growth Factor Signaling in Embryos. Taylor and Francis Books; 2006b. pp. 177–262.
-
- Albrieux M, Villaz M. Bilateral asymmetry of the inositol trisphosphate-mediated calcium signaling in two-cell ascidian embryos. Biology of the Cell. 2000;92:277–84. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
