

22-Nucleotide RNAs trigger secondary siRNA biogenesis in plants
- PMID: 20643946
- PMCID: PMC2930544
- DOI: 10.1073/pnas.1001738107
22-Nucleotide RNAs trigger secondary siRNA biogenesis in plants
Abstract
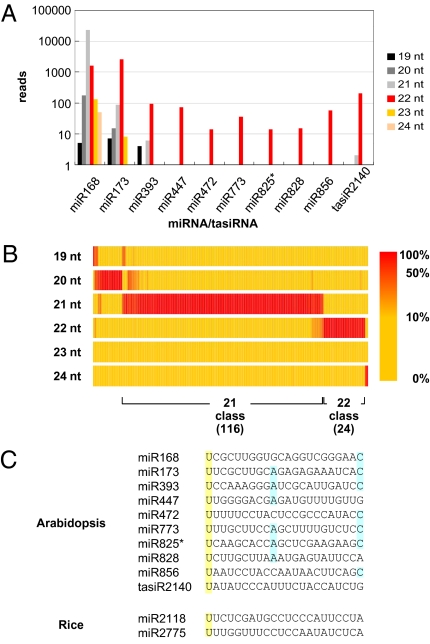
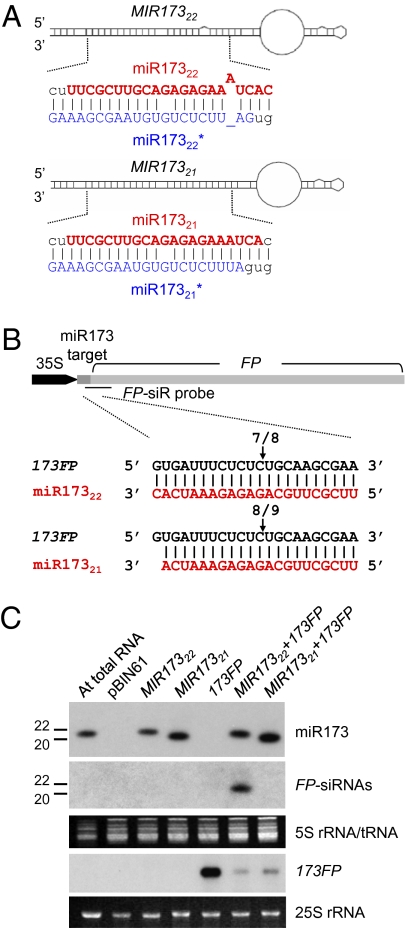
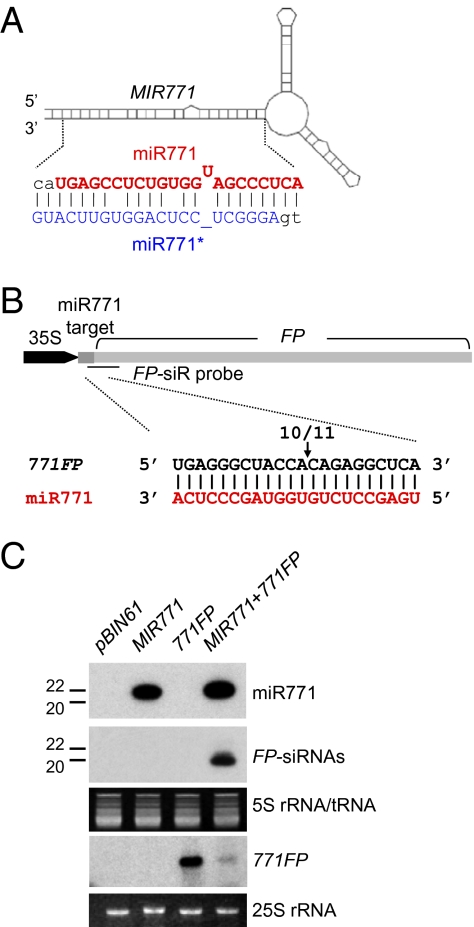
The effect of RNA silencing in plants can be amplified if the production of secondary small interfering RNAs (siRNAs) is triggered by the interaction of microRNAs (miRNAs) or siRNAs with a long target RNA. miRNA and siRNA interactions are not all equivalent, however; most of them do not trigger secondary siRNA production. Here we use bioinformatics to show that the secondary siRNA triggers are miRNAs and transacting siRNAs of 22 nt, rather than the more typical 21-nt length. Agrobacterium-mediated transient expression in Nicotiana benthamiana confirms that the siRNA-initiating miRNAs, miR173 and miR828, are effective as triggers only if expressed in a 22-nt form and, conversely, that increasing the length of miR319 from 21 to 22 nt converts it to an siRNA trigger. We also predicted and validated that the 22-nt miR771 is a secondary siRNA trigger. Our data demonstrate that the function of small RNAs is influenced by size, and that a length of 22 nt facilitates the triggering of secondary siRNA production.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Comment in
-
RNA silencing amplification in plants: size matters.Proc Natl Acad Sci U S A. 2010 Aug 24;107(34):14945-6. doi: 10.1073/pnas.1009416107. Epub 2010 Aug 13. Proc Natl Acad Sci U S A. 2010. PMID: 20709960 Free PMC article. No abstract available.
References
-
- Voinnet O. Use, tolerance and avoidance of amplified RNA silencing by plants. Trends Plant Sci. 2008;13:317–328. - PubMed
-
- Vaucheret H. MicroRNA-dependent trans-acting siRNA production. Sci STKE. 2005;2005:pe43. - PubMed
-
- Allen E, Xie Z, Gustafson AM, Carrington JC. MicroRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell. 2005;121:207–221. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
